“霍奇金-赫胥黎模型”的版本间的差异
| 第12行: | 第12行: | ||
[[Alan Hodgkin]] and [[Andrew Huxley]] described the model in 1952 to explain the ionic mechanisms underlying the initiation and propagation of action potentials in the [[squid giant axon]].<ref name=HH>{{cite journal | vauthors = Hodgkin AL, Huxley AF | title = A quantitative description of membrane current and its application to conduction and excitation in nerve | journal = The Journal of Physiology | volume = 117 | issue = 4 | pages = 500–44 | date = August 1952 | pmid = 12991237 | pmc = 1392413 | doi = 10.1113/jphysiol.1952.sp004764 }}</ref> They received the 1963 [[Nobel Prize in Physiology or Medicine]] for this work. | [[Alan Hodgkin]] and [[Andrew Huxley]] described the model in 1952 to explain the ionic mechanisms underlying the initiation and propagation of action potentials in the [[squid giant axon]].<ref name=HH>{{cite journal | vauthors = Hodgkin AL, Huxley AF | title = A quantitative description of membrane current and its application to conduction and excitation in nerve | journal = The Journal of Physiology | volume = 117 | issue = 4 | pages = 500–44 | date = August 1952 | pmid = 12991237 | pmc = 1392413 | doi = 10.1113/jphysiol.1952.sp004764 }}</ref> They received the 1963 [[Nobel Prize in Physiology or Medicine]] for this work. | ||
| − | 1952年,[[Alan Hodgkin]] 和 [[Andrew Huxley]] 描述了这个模型,来解释[[乌贼巨大轴突]]中动作电位的产生和传导的离子机制。<ref name= | + | 1952年,[[Alan Hodgkin]] 和 [[Andrew Huxley]] 描述了这个模型,来解释[[乌贼巨大轴突]]中动作电位的产生和传导的离子机制。<ref name=HH>{{cite journal | vauthors = Hodgkin AL, Huxley AF | title = A quantitative description of membrane current and its application to conduction and excitation in nerve | journal = The Journal of Physiology | volume = 117 | issue = 4 | pages = 500–44 | date = August 1952 | pmid = 12991237 | pmc = 1392413 | doi = 10.1113/jphysiol.1952.sp004764 }}</ref>他们因为这项工作获得了1963年的[[诺贝尔生理学或医学奖]]。 |
==基本组成部分== | ==基本组成部分== | ||
| − | The typical Hodgkin–Huxley model treats each component of an excitable cell as an electrical element (as shown in the figure). The [[lipid bilayer]] is represented as a [[capacitance]] (C<SUB>m</SUB>). [[Voltage-gated ion channel]]s are represented by [[electrical conductance]]s (''g''<SUB>''n''</SUB>, where ''n'' is the specific ion channel) that depend on both voltage and time. [[Leak channel]]s are represented by linear conductances (''g''<SUB>''L''</SUB>). The [[electrochemical gradient]]s driving the flow of ions are represented by [[voltage source]]s (''E''<SUB>''n''</SUB>) whose [[voltage]]s are determined by the ratio of the intra- and extracellular concentrations of the ionic species of interest. Finally, [[Ion pump (biology)|ion pumps]] are represented by [[current sources]] (''I''<SUB>''p''</SUB>). | + | The typical Hodgkin–Huxley model treats each component of an excitable cell as an electrical element (as shown in the figure). The [[lipid bilayer]] is represented as a [[capacitance]] (C<SUB>m</SUB>). [[Voltage-gated ion channel]]s are represented by [[electrical conductance]]s (''g''<SUB>''n''</SUB>, where ''n'' is the specific ion channel) that depend on both voltage and time. [[Leak channel]]s are represented by linear conductances (''g''<SUB>''L''</SUB>). The [[electrochemical gradient]]s driving the flow of ions are represented by [[voltage source]]s (''E''<SUB>''n''</SUB>) whose [[voltage]]s are determined by the ratio of the intra- and extracellular concentrations of the ionic species of interest. Finally, [[Ion pump (biology)|ion pumps]] are represented by [[current sources]] (''I''<SUB>''p''</SUB>). The [[membrane potential]] is denoted by ''V<SUB>m</SUB>''. |
经典的 Hodgkin-Huxley 模型将可兴奋细胞的每个部分都当作电路元件来处理(如图所示)。[[磷脂双分子层]]表示为电容(<math>C_m</math>)。[[电压门控离子通道]]表示为[[电导]](<math>g_n</math>,其中 n 是特定的离子通道),它依赖于电压和时间。[[漏通道]]表示为线性电导(<math>g_L</math>)。驱使离子流动的[[电化学梯度]]表示为[[电压源]](<math>E_n</math>),电压源的[[电压]]取决于相关离子种类在细胞内和细胞外的浓度的比值。最后,[[离子泵]]表示为[[电流源]](<math>i_p</math>)。[[膜电位]]表示为<math>V_m</math>。 | 经典的 Hodgkin-Huxley 模型将可兴奋细胞的每个部分都当作电路元件来处理(如图所示)。[[磷脂双分子层]]表示为电容(<math>C_m</math>)。[[电压门控离子通道]]表示为[[电导]](<math>g_n</math>,其中 n 是特定的离子通道),它依赖于电压和时间。[[漏通道]]表示为线性电导(<math>g_L</math>)。驱使离子流动的[[电化学梯度]]表示为[[电压源]](<math>E_n</math>),电压源的[[电压]]取决于相关离子种类在细胞内和细胞外的浓度的比值。最后,[[离子泵]]表示为[[电流源]](<math>i_p</math>)。[[膜电位]]表示为<math>V_m</math>。 | ||
| 第47行: | 第47行: | ||
In voltage-gated ion channels, the channel conductance <math>g_l</math> is a function of both time and voltage (<math>g_n(t,V)</math> in the figure), while in leak channels <math>g_l</math> is a constant (<math>g_L</math> in the figure). The current generated by ion pumps is dependent on the ionic species specific to that pump. The following sections will describe these formulations in more detail. | In voltage-gated ion channels, the channel conductance <math>g_l</math> is a function of both time and voltage (<math>g_n(t,V)</math> in the figure), while in leak channels <math>g_l</math> is a constant (<math>g_L</math> in the figure). The current generated by ion pumps is dependent on the ionic species specific to that pump. The following sections will describe these formulations in more detail. | ||
| − | 在电压门控离子通道中,通道电导<math>g_l</math>是时间和电压(图中为 <math>g_n(t,V)</math>)的函数,而在漏通道中,<math>g_l</math>是常数(图中为<math>g_L</math> | + | 在电压门控离子通道中,通道电导<math>g_l</math>是时间和电压(图中为 <math>g_n(t,V)</math>)的函数,而在漏通道中,<math>g_l</math>是常数(图中为<math>g_L</math>)。由离子泵产生的电流取决于离子泵特定的离子种类。以下各节将更详细地描述这些公式。 |
| − | |||
| − | |||
===电压门控离子通道=== | ===电压门控离子通道=== | ||
Using a series of [[voltage clamp]] experiments and by varying extracellular sodium and potassium concentrations, Hodgkin and Huxley developed a model in which the properties of an excitable cell are described by a set of four [[ordinary differential equation]]s.<ref name="HH"/> Together with the equation for the total current mentioned above, these are: | Using a series of [[voltage clamp]] experiments and by varying extracellular sodium and potassium concentrations, Hodgkin and Huxley developed a model in which the properties of an excitable cell are described by a set of four [[ordinary differential equation]]s.<ref name="HH"/> Together with the equation for the total current mentioned above, these are: | ||
| − | 通过改变细胞外钠离子和钾离子的浓度,进行一系列的[[电压钳]] | + | 通过改变细胞外钠离子和钾离子的浓度,进行一系列的[[电压钳]]实验,Hodgkin和Huxley建立了一个由四个[[常微分方程]]描述的可兴奋细胞特性的模型。<ref name="HH"/>加上上述总电流的方程,这些方程为: |
: <math>I = C_m\frac{{\mathrm d} V_m}{{\mathrm d} t} + \bar{g}_\text{K}n^4(V_m - V_K) + \bar{g}_\text{Na}m^3h(V_m - V_{Na}) + \bar{g}_l(V_m - V_l),</math> | : <math>I = C_m\frac{{\mathrm d} V_m}{{\mathrm d} t} + \bar{g}_\text{K}n^4(V_m - V_K) + \bar{g}_\text{Na}m^3h(V_m - V_{Na}) + \bar{g}_l(V_m - V_l),</math> | ||
| 第66行: | 第64行: | ||
where ''I'' is the current per unit area, and <math>\alpha_i </math> and <math>\beta_i </math> are rate constants for the ''i''-th ion channel, which depend on voltage but not time. <math>\bar{g}_n</math> is the maximal value of the conductance. ''n'', ''m'', and ''h'' are dimensionless quantities between 0 and 1 that are associated with potassium channel activation, sodium channel activation, and sodium channel inactivation, respectively. For <math> p = (n, m, h)</math>, <math> \alpha_p </math> and <math> \beta_p </math> take the form | where ''I'' is the current per unit area, and <math>\alpha_i </math> and <math>\beta_i </math> are rate constants for the ''i''-th ion channel, which depend on voltage but not time. <math>\bar{g}_n</math> is the maximal value of the conductance. ''n'', ''m'', and ''h'' are dimensionless quantities between 0 and 1 that are associated with potassium channel activation, sodium channel activation, and sodium channel inactivation, respectively. For <math> p = (n, m, h)</math>, <math> \alpha_p </math> and <math> \beta_p </math> take the form | ||
| − | 其中<math>I</math>是单位面积的电流,而<math>\alpha_i</math>和<math>\beta_i</math>是第<math>i</math>个离子通道的速率常数,它取决于电压而非时间。<math>\bar{g}_n</math>是电导的最大值。<math>N</math>、 <math>m</math> 和 <math>h</math> 是0和1之间的无量纲量,分别与钾通道激活、钠通道激活和钠通道失活有关。对于<math>p = (n,m,h)</math>,<math>\alpha_p</math>和<math>\beta_p</math>的形式是 | + | 其中<math>I</math>是单位面积的电流,而<math>\alpha_i</math>和<math>\beta_i</math>是第<math>i</math>个离子通道的速率常数,它取决于电压而非时间。<math>\bar{g}_n</math>是电导的最大值。<math>N</math>、<math>m</math>和<math>h</math>是0和1之间的无量纲量,分别与钾通道激活、钠通道激活和钠通道失活有关。对于<math>p = (n,m,h)</math>,<math>\alpha_p</math>和<math>\beta_p</math>的形式是 |
: <math>\alpha_p(V_m) = p_\infty(V_m)/\tau_p</math> | : <math>\alpha_p(V_m) = p_\infty(V_m)/\tau_p</math> | ||
| 第79行: | 第77行: | ||
where <math> V = V_{rest} - V_m </math> denotes the negative depolarization in mV. | where <math> V = V_{rest} - V_m </math> denotes the negative depolarization in mV. | ||
| + | <math>p_\infty</math> 和 <math>(1-p_\infty)</math>分别是激活和失活的稳态值,通常用[[玻尔兹曼方程]]表示为<math>V_m</math>的函数。 | ||
| − | |||
在 Hodgkin 和 Huxley 的原始论文<ref name="HH"/>中,<math>\alpha</math>和<math>\beta</math>函数如下给出 | 在 Hodgkin 和 Huxley 的原始论文<ref name="HH"/>中,<math>\alpha</math>和<math>\beta</math>函数如下给出 | ||
| 第89行: | 第87行: | ||
\end{array} </math> | \end{array} </math> | ||
| − | + | 其中<math> V = V_{rest} - V_m </math> <font color = "#32CD32">表示负去极化</font>,单位为mV。 | |
| − | 其中<math> V = V_{rest} - V_m </math | ||
| − | |||
While in many current software programs,<ref>Nelson ME (2005) [http://nelson.beckman.illinois.edu/courses/physl317/part1/Lec3_HHsection.pdf Electrophysiological Models In: Databasing the Brain: From Data to Knowledge.] (S. Koslow and S. Subramaniam, eds.) Wiley, New York, pp. 285–301</ref> Hodgkin–Huxley type models generalize <math> \alpha </math> and <math> \beta </math> to | While in many current software programs,<ref>Nelson ME (2005) [http://nelson.beckman.illinois.edu/courses/physl317/part1/Lec3_HHsection.pdf Electrophysiological Models In: Databasing the Brain: From Data to Knowledge.] (S. Koslow and S. Subramaniam, eds.) Wiley, New York, pp. 285–301</ref> Hodgkin–Huxley type models generalize <math> \alpha </math> and <math> \beta </math> to | ||
| − | + | 在当前的软件程序中<ref>Nelson ME (2005) [http://nelson.beckman.illinois.edu/courses/physl317/part1/Lec3_HHsection.pdf Electrophysiological Models In: Databasing the Brain: From Data to Knowledge.] (S. Koslow and S. Subramaniam, eds.) Wiley, New York, pp. 285–301</ref>,霍奇金-赫胥黎类模型将<math>\alpha</math>和><math>\beta</math>归纳为 | |
:<math> \frac{A_p(V_m-B_p)}{\exp\big(\frac{V_m-B_p}{C_p}\big)-D_p} </math> | :<math> \frac{A_p(V_m-B_p)}{\exp\big(\frac{V_m-B_p}{C_p}\big)-D_p} </math> | ||
| 第101行: | 第97行: | ||
In order to characterize voltage-gated channels, the equations are fit to voltage clamp data. For a derivation of the Hodgkin–Huxley equations under voltage-clamp, see.<ref name="JohnstonAndWu">{{cite book|last1=Gray|first1=Daniel Johnston|first2=Samuel Miao-Sin|last2=Wu | name-list-style = vanc |title=Foundations of cellular neurophysiology|year=1997|publisher=MIT Press|location=Cambridge, Massachusetts [u.a.]|isbn=978-0-262-10053-3|edition=3rd}}</ref> Briefly, when the membrane potential is held at a constant value (i.e., voltage-clamp), for each value of the membrane potential the nonlinear gating equations reduce to equations of the form: | In order to characterize voltage-gated channels, the equations are fit to voltage clamp data. For a derivation of the Hodgkin–Huxley equations under voltage-clamp, see.<ref name="JohnstonAndWu">{{cite book|last1=Gray|first1=Daniel Johnston|first2=Samuel Miao-Sin|last2=Wu | name-list-style = vanc |title=Foundations of cellular neurophysiology|year=1997|publisher=MIT Press|location=Cambridge, Massachusetts [u.a.]|isbn=978-0-262-10053-3|edition=3rd}}</ref> Briefly, when the membrane potential is held at a constant value (i.e., voltage-clamp), for each value of the membrane potential the nonlinear gating equations reduce to equations of the form: | ||
| − | 为了刻画电压门控通道,'''<font color="#32CD32"> | + | 为了刻画电压门控通道,'''<font color="#32CD32">该方程拟合自电压钳数据</font>'''。关于电压钳下Hodgkin-Huxley方程的推导,请参阅<ref name="JohnstonAndWu">{{cite book|last1=Gray|first1=Daniel Johnston|first2=Samuel Miao-Sin|last2=Wu | name-list-style = vanc |title=Foundations of cellular neurophysiology|year=1997|publisher=MIT Press|location=Cambridge, Massachusetts [u.a.]|isbn=978-0-262-10053-3|edition=3rd}}</ref>。简单来说,当膜电位保持为一个恒定值(即电压钳取值)时,对于膜电位的每个值,非线性门控方程可以归结为以下形式的方程: |
: <math>m(t) = m_{0} - [ (m_{0}-m_{\infty})(1 - e^{-t/\tau_m})]\, </math> | : <math>m(t) = m_{0} - [ (m_{0}-m_{\infty})(1 - e^{-t/\tau_m})]\, </math> | ||
| 第119行: | 第115行: | ||
In order to arrive at the complete solution for a propagated action potential, one must write the current term ''I'' on the left-hand side of the first differential equation in terms of ''V'', so that the equation becomes an equation for voltage alone. The relation between ''I'' and ''V'' can be derived from [[cable theory]] and is given by | In order to arrive at the complete solution for a propagated action potential, one must write the current term ''I'' on the left-hand side of the first differential equation in terms of ''V'', so that the equation becomes an equation for voltage alone. The relation between ''I'' and ''V'' can be derived from [[cable theory]] and is given by | ||
| − | 为了得到传导的动作电位的完全解, | + | 为了得到传导的动作电位的完全解,<font color="#32CD32"> 必须将第一个微分方程左侧的电流项 <math>I</math>写为<math>V</math>的形式</font>,使方程成为单独的电压方程。<math>I</math>和<math>V</math>之间的关系可以从[[电缆理论]]中推导出来,即 |
: <math>I = \frac{a}{2R}\frac{\partial^2V}{\partial x^2}, </math> | : <math>I = \frac{a}{2R}\frac{\partial^2V}{\partial x^2}, </math> | ||
| 第125行: | 第121行: | ||
where ''a'' is the radius of the [[axon]], ''R'' is the [[Resistivity|specific resistance]] of the [[axoplasm]], and ''x'' is the position along the nerve fiber. Substitution of this expression for ''I'' transforms the original set of equations into a set of [[partial differential equation]]s, because the voltage becomes a function of both ''x'' and ''t''. | where ''a'' is the radius of the [[axon]], ''R'' is the [[Resistivity|specific resistance]] of the [[axoplasm]], and ''x'' is the position along the nerve fiber. Substitution of this expression for ''I'' transforms the original set of equations into a set of [[partial differential equation]]s, because the voltage becomes a function of both ''x'' and ''t''. | ||
| − | 其中<math>a</math>是[[轴突]]的半径,<math>R</math> 是[[轴浆]]的[[比电阻]],<math>x</math>是沿神经纤维的位置。用这个表达式代替<math>I</math>,将原来的方程组转变为一组[[偏微分方程]],因为电压变为<math>x</math>和<math>t</math>的函数。 | + | 其中<math>a</math>是[[轴突]]的半径,<math>R</math>是[[轴浆]]的[[比电阻]],<math>x</math>是沿神经纤维的位置。用这个表达式代替<math>I</math>,将原来的方程组转变为一组[[偏微分方程]],因为电压变为<math>x</math>和<math>t</math>的函数。 |
The [[Levenberg–Marquardt algorithm]] is often used to fit these equations to voltage-clamp data.<ref>{{cite book |title=New Ecoinformatics Tools in Environmental Science : Applications and Decision-making |first1=Vladimir F. |last1=Krapivin |first2=Costas A. |last2=Varotsos |first3=Vladimir Yu. |last3=Soldatov |year=2015 |pages=37–38 |publisher=Springer |isbn=9783319139784 |url=https://www.google.com/books/edition/New_Ecoinformatics_Tools_in_Environmenta/bWpnBgAAQBAJ?hl=en&gbpv=1&pg=PA37 }}</ref> | The [[Levenberg–Marquardt algorithm]] is often used to fit these equations to voltage-clamp data.<ref>{{cite book |title=New Ecoinformatics Tools in Environmental Science : Applications and Decision-making |first1=Vladimir F. |last1=Krapivin |first2=Costas A. |last2=Varotsos |first3=Vladimir Yu. |last3=Soldatov |year=2015 |pages=37–38 |publisher=Springer |isbn=9783319139784 |url=https://www.google.com/books/edition/New_Ecoinformatics_Tools_in_Environmenta/bWpnBgAAQBAJ?hl=en&gbpv=1&pg=PA37 }}</ref> | ||
| 第133行: | 第129行: | ||
While the original experiments treated only sodium and potassium channels, the Hodgkin–Huxley model can also be extended to account for other species of [[ion channel]]s. | While the original experiments treated only sodium and potassium channels, the Hodgkin–Huxley model can also be extended to account for other species of [[ion channel]]s. | ||
| − | 虽然最初的实验只处理钠和钾通道,Hodgkin- | + | 虽然最初的实验只处理钠和钾通道,Hodgkin-Huxley模型也可以扩展到其他种类的离子通道。 |
===漏通道=== | ===漏通道=== | ||
Leak channels account for the natural permeability of the membrane to ions and take the form of the equation for voltage-gated channels, where the conductance <math>g_{leak}</math> is a constant. Thus, the leak current due to passive leak ion channels in the Hodgkin-Huxley formalism is <math>I_l=g_{leak}(V-V_{leak})</math>. | Leak channels account for the natural permeability of the membrane to ions and take the form of the equation for voltage-gated channels, where the conductance <math>g_{leak}</math> is a constant. Thus, the leak current due to passive leak ion channels in the Hodgkin-Huxley formalism is <math>I_l=g_{leak}(V-V_{leak})</math>. | ||
| − | 漏通道解释了膜对离子的天然渗透性,其形式为电压门控通道方程,其中电导<math>g_{leak}</math> | + | 漏通道解释了膜对离子的天然渗透性,其形式为电压门控通道方程,其中电导<math>g_{leak}</math>为常数。因此,在Hodgkin-Huxley公式中,被动漏离子通道引起的漏电流为<math>I_l=g_{leak}(V-V_{leak})</math>。 |
| − | |||
===泵和交换器=== | ===泵和交换器=== | ||
| 第145行: | 第140行: | ||
The membrane potential depends upon the maintenance of ionic concentration gradients across it. The maintenance of these concentration gradients requires active transport of ionic species. The sodium-potassium and sodium-calcium exchangers are the best known of these. Some of the basic properties of the Na/Ca exchanger have already been well-established: the stoichiometry of exchange is 3 Na<SUP>+</SUP>: 1 Ca<SUP>2+</SUP> and the exchanger is electrogenic and voltage-sensitive. The Na/K exchanger has also been described in detail, with a 3 Na<SUP>+</SUP>: 2 K<SUP>+</SUP> stoichiometry.<ref name="Rakowski_1989">{{cite journal | vauthors = Rakowski RF, Gadsby DC, De Weer P | title = Stoichiometry and voltage dependence of the sodium pump in voltage-clamped, internally dialyzed squid giant axon | journal = The Journal of General Physiology | volume = 93 | issue = 5 | pages = 903–41 | date = May 1989 | pmid = 2544655 | doi = 10.1085/jgp.93.5.903 | pmc=2216238}}</ref><ref name="Hille">{{cite book|last=Hille|first=Bertil | name-list-style = vanc | title=Ion channels of excitable membranes|year=2001|publisher=Sinauer|location=Sunderland, Massachusetts|isbn=978-0-87893-321-1 | edition = 3rd }}</ref> | The membrane potential depends upon the maintenance of ionic concentration gradients across it. The maintenance of these concentration gradients requires active transport of ionic species. The sodium-potassium and sodium-calcium exchangers are the best known of these. Some of the basic properties of the Na/Ca exchanger have already been well-established: the stoichiometry of exchange is 3 Na<SUP>+</SUP>: 1 Ca<SUP>2+</SUP> and the exchanger is electrogenic and voltage-sensitive. The Na/K exchanger has also been described in detail, with a 3 Na<SUP>+</SUP>: 2 K<SUP>+</SUP> stoichiometry.<ref name="Rakowski_1989">{{cite journal | vauthors = Rakowski RF, Gadsby DC, De Weer P | title = Stoichiometry and voltage dependence of the sodium pump in voltage-clamped, internally dialyzed squid giant axon | journal = The Journal of General Physiology | volume = 93 | issue = 5 | pages = 903–41 | date = May 1989 | pmid = 2544655 | doi = 10.1085/jgp.93.5.903 | pmc=2216238}}</ref><ref name="Hille">{{cite book|last=Hille|first=Bertil | name-list-style = vanc | title=Ion channels of excitable membranes|year=2001|publisher=Sinauer|location=Sunderland, Massachusetts|isbn=978-0-87893-321-1 | edition = 3rd }}</ref> | ||
| − | 膜电位取决于其跨膜离子浓度梯度的保持。维持这些浓度梯度需要这几种离子的主动运输。其中钠钾交换器和钠钙交换器最为著名。钠钙交换器的一些基本性质已得到公认: | + | 膜电位取决于其跨膜离子浓度梯度的保持。维持这些浓度梯度需要这几种离子的主动运输。其中钠钾交换器和钠钙交换器最为著名。钠钙交换器的一些基本性质已得到公认: 交换的化学计量比为 3 Na<SUP>+</SUP>: 1 Ca<SUP>2+</SUP>,且具有生电性和电压敏感性。文献中还详细描述了Na/K交换器,它具有 3 Na<SUP>+</SUP>: 2 K<SUP>+</SUP>的化学计量比。<ref name="Rakowski_1989">{{cite journal | vauthors = Rakowski RF, Gadsby DC, De Weer P | title = Stoichiometry and voltage dependence of the sodium pump in voltage-clamped, internally dialyzed squid giant axon | journal = The Journal of General Physiology | volume = 93 | issue = 5 | pages = 903–41 | date = May 1989 | pmid = 2544655 | doi = 10.1085/jgp.93.5.903 | pmc=2216238}}</ref><ref name="Hille">{{cite book|last=Hille|first=Bertil | name-list-style = vanc | title=Ion channels of excitable membranes|year=2001|publisher=Sinauer|location=Sunderland, Massachusetts|isbn=978-0-87893-321-1 | edition = 3rd }}</ref> |
==数学性质== | ==数学性质== | ||
| 第192行: | 第187行: | ||
霍奇金-赫胥黎模型被认为是20世纪生物物理学的伟大成就之一。尽管如此,现代 Hodgkin-Huxley 型模型已经在几个重要方面得到了扩展: | 霍奇金-赫胥黎模型被认为是20世纪生物物理学的伟大成就之一。尽管如此,现代 Hodgkin-Huxley 型模型已经在几个重要方面得到了扩展: | ||
* 根据实验数据引入了额外的离子通道群。 | * 根据实验数据引入了额外的离子通道群。 | ||
| − | * 对Hodgkin-Huxley 模型加以修正,加入了[[过渡态理论]],并产生了[[热力学]] Hodgkin-Huxley 模型。<ref>{{cite journal |last=Forrest |first=M. D. |title=Can the Thermodynamic Hodgkin–Huxley Model of Voltage-Dependent Conductance Extrapolate for Temperature? |journal=Computation |volume=2 |issue=2 |pages=47–60 |date=May 2014 |doi=10.3390/computation2020047|url=http://wrap.warwick.ac.uk/60495/1/WRAP_computation-02-00047.pdf |doi-access=free }}</ref> | + | * 对Hodgkin-Huxley 模型加以修正,加入了[[过渡态理论]],并产生了[[热力学]] Hodgkin-Huxley 模型。<ref>{{cite journal |last=Forrest |first=M. D. |title=Can the Thermodynamic Hodgkin–Huxley Model of Voltage-Dependent Conductance Extrapolate for Temperature? |journal=Computation |volume=2 |issue=2 |pages=47–60 |date=May 2014 |doi=10.3390/computation2020047|url=http://wrap.warwick.ac.uk/60495/1/WRAP_computation-02-00047.pdf |doi-access=free }}</ref> |
* 通常基于显微镜数据,模型通常包含[[树突]]和[[轴突]]的高度复杂的几何形状。 | * 通常基于显微镜数据,模型通常包含[[树突]]和[[轴突]]的高度复杂的几何形状。 | ||
* 离子通道行为的[[随机]]模型,导致随机混合系统。<ref name=stochastic>{{cite journal |last1=Pakdaman |first1=K. |last2=Thieullen |first2=M. |first3=G. |last3=Wainrib |title=Fluid limit theorems for stochastic hybrid systems with applications to neuron models |year=2010 |journal=Adv. Appl. Probab. |volume=42 |issue=3 |pages=761–794 |doi=10.1239/aap/1282924062 |arxiv=1001.2474 |bibcode=2010arXiv1001.2474P|s2cid=18894661 }}</ref> | * 离子通道行为的[[随机]]模型,导致随机混合系统。<ref name=stochastic>{{cite journal |last1=Pakdaman |first1=K. |last2=Thieullen |first2=M. |first3=G. |last3=Wainrib |title=Fluid limit theorems for stochastic hybrid systems with applications to neuron models |year=2010 |journal=Adv. Appl. Probab. |volume=42 |issue=3 |pages=761–794 |doi=10.1239/aap/1282924062 |arxiv=1001.2474 |bibcode=2010arXiv1001.2474P|s2cid=18894661 }}</ref> | ||
2022年3月30日 (三) 06:39的版本
此词条由神经动力学读书会词条梳理志愿者安贞桦翻译审校,未经专家审核,带来阅读不便,请见谅。

{kind=link}
The Hodgkin–Huxley model, or conductance-based model, is a mathematical model that describes how action potentials in neurons are initiated and propagated. It is a set of nonlinear differential equations that approximates the electrical characteristics of excitable cells such as neurons and cardiac myocytes. It is a continuous-time dynamical system.
霍奇金-赫胥黎模型Hodgkin-Huxley model,或者说基于电导的模型conductance-based model,是一个描述神经元中动作电位如何产生和传导的数学模型。它是一组非线性微分方程,用于近似可兴奋细胞(如神经元和心肌细胞)的电学特性。它是一个时间连续的动力系统。
Alan Hodgkin and Andrew Huxley described the model in 1952 to explain the ionic mechanisms underlying the initiation and propagation of action potentials in the squid giant axon.[1] They received the 1963 Nobel Prize in Physiology or Medicine for this work.
1952年,Alan Hodgkin 和 Andrew Huxley 描述了这个模型,来解释乌贼巨大轴突中动作电位的产生和传导的离子机制。[1]他们因为这项工作获得了1963年的诺贝尔生理学或医学奖。
基本组成部分
The typical Hodgkin–Huxley model treats each component of an excitable cell as an electrical element (as shown in the figure). The lipid bilayer is represented as a capacitance (Cm). Voltage-gated ion channels are represented by electrical conductances (gn, where n is the specific ion channel) that depend on both voltage and time. Leak channels are represented by linear conductances (gL). The electrochemical gradients driving the flow of ions are represented by voltage sources (En) whose voltages are determined by the ratio of the intra- and extracellular concentrations of the ionic species of interest. Finally, ion pumps are represented by current sources (Ip). The membrane potential is denoted by Vm.
经典的 Hodgkin-Huxley 模型将可兴奋细胞的每个部分都当作电路元件来处理(如图所示)。磷脂双分子层表示为电容([math]\displaystyle{ C_m }[/math])。电压门控离子通道表示为电导([math]\displaystyle{ g_n }[/math],其中 n 是特定的离子通道),它依赖于电压和时间。漏通道表示为线性电导([math]\displaystyle{ g_L }[/math])。驱使离子流动的电化学梯度表示为电压源([math]\displaystyle{ E_n }[/math]),电压源的电压取决于相关离子种类在细胞内和细胞外的浓度的比值。最后,离子泵表示为电流源([math]\displaystyle{ i_p }[/math])。膜电位表示为[math]\displaystyle{ V_m }[/math]。
Mathematically, the current flowing through the lipid bilayer is written as
数学上,流过磷脂双分子层的电流写为
- [math]\displaystyle{ I_c = C_m\frac{{\mathrm d} V_m}{{\mathrm d} t} }[/math]
and the current through a given ion channel is the product
流过给定离子通道的电流为乘积
- [math]\displaystyle{ I_i = {g_i}(V_m - V_i) \; }[/math]
where [math]\displaystyle{ V_i }[/math] is the reversal potential of the 第[math]\displaystyle{ i }[/math]个 ion channel. Thus, for a cell with sodium and potassium channels, the total current through the membrane is given by:
其中 [math]\displaystyle{ V_i }[/math] 是第[math]\displaystyle{ i }[/math]个离子通道的反转电位。因此,对于具有钠和钾离子通道的细胞,通过细胞膜的总电流为:
- [math]\displaystyle{ I = C_m\frac{{\mathrm d} V_m}{{\mathrm d} t} + g_K(V_m - V_K) + g_{Na}(V_m - V_{Na}) + g_l(V_m - V_l) }[/math]
where I is the total membrane current per unit area, Cm is the membrane capacitance per unit area, gK and gNa are the potassium and sodium conductances per unit area, respectively, VK and VNa are the potassium and sodium reversal potentials, respectively, and gl and Vl are the leak conductance per unit area and leak reversal potential, respectively. The time dependent elements of this equation are Vm, gNa, and gK, where the last two conductances depend explicitly on voltage as well.
其中[math]\displaystyle{ I }[/math]为单位面积的总膜电流,[math]\displaystyle{ C_m }[/math]为单位面积的膜电容,[math]\displaystyle{ g_K }[/math]和[math]\displaystyle{ g_{Na} }[/math]分别为单位面积的钾和钠的电导,[math]\displaystyle{ V_K }[/math]和[math]\displaystyle{ V_{Na} }[/math]分别为钾和钠的反转电位,[math]\displaystyle{ g_l }[/math]和[math]\displaystyle{ V_l }[/math]分别为单位面积的漏电导和漏反转电位。这个方程中对时间依赖的项为[math]\displaystyle{ V_m }[/math]、[math]\displaystyle{ g_{Na} }[/math]和[math]\displaystyle{ g_K }[/math],其中最后两个电导项也明确地取决于电压。
离子电流的刻画
In voltage-gated ion channels, the channel conductance [math]\displaystyle{ g_l }[/math] is a function of both time and voltage ([math]\displaystyle{ g_n(t,V) }[/math] in the figure), while in leak channels [math]\displaystyle{ g_l }[/math] is a constant ([math]\displaystyle{ g_L }[/math] in the figure). The current generated by ion pumps is dependent on the ionic species specific to that pump. The following sections will describe these formulations in more detail.
在电压门控离子通道中,通道电导[math]\displaystyle{ g_l }[/math]是时间和电压(图中为 [math]\displaystyle{ g_n(t,V) }[/math])的函数,而在漏通道中,[math]\displaystyle{ g_l }[/math]是常数(图中为[math]\displaystyle{ g_L }[/math])。由离子泵产生的电流取决于离子泵特定的离子种类。以下各节将更详细地描述这些公式。
电压门控离子通道
Using a series of voltage clamp experiments and by varying extracellular sodium and potassium concentrations, Hodgkin and Huxley developed a model in which the properties of an excitable cell are described by a set of four ordinary differential equations.[1] Together with the equation for the total current mentioned above, these are:
通过改变细胞外钠离子和钾离子的浓度,进行一系列的电压钳实验,Hodgkin和Huxley建立了一个由四个常微分方程描述的可兴奋细胞特性的模型。[1]加上上述总电流的方程,这些方程为:
- [math]\displaystyle{ I = C_m\frac{{\mathrm d} V_m}{{\mathrm d} t} + \bar{g}_\text{K}n^4(V_m - V_K) + \bar{g}_\text{Na}m^3h(V_m - V_{Na}) + \bar{g}_l(V_m - V_l), }[/math]
- [math]\displaystyle{ \frac{dn}{dt} = \alpha_n(V_m)(1 - n) - \beta_n(V_m) n }[/math]
- [math]\displaystyle{ \frac{dm}{dt} = \alpha_m(V_m)(1 - m) - \beta_m(V_m) m }[/math]
- [math]\displaystyle{ \frac{dh}{dt} = \alpha_h(V_m)(1 - h) - \beta_h(V_m) h }[/math]
where I is the current per unit area, and [math]\displaystyle{ \alpha_i }[/math] and [math]\displaystyle{ \beta_i }[/math] are rate constants for the i-th ion channel, which depend on voltage but not time. [math]\displaystyle{ \bar{g}_n }[/math] is the maximal value of the conductance. n, m, and h are dimensionless quantities between 0 and 1 that are associated with potassium channel activation, sodium channel activation, and sodium channel inactivation, respectively. For [math]\displaystyle{ p = (n, m, h) }[/math], [math]\displaystyle{ \alpha_p }[/math] and [math]\displaystyle{ \beta_p }[/math] take the form
其中[math]\displaystyle{ I }[/math]是单位面积的电流,而[math]\displaystyle{ \alpha_i }[/math]和[math]\displaystyle{ \beta_i }[/math]是第[math]\displaystyle{ i }[/math]个离子通道的速率常数,它取决于电压而非时间。[math]\displaystyle{ \bar{g}_n }[/math]是电导的最大值。[math]\displaystyle{ N }[/math]、[math]\displaystyle{ m }[/math]和[math]\displaystyle{ h }[/math]是0和1之间的无量纲量,分别与钾通道激活、钠通道激活和钠通道失活有关。对于[math]\displaystyle{ p = (n,m,h) }[/math],[math]\displaystyle{ \alpha_p }[/math]和[math]\displaystyle{ \beta_p }[/math]的形式是
- [math]\displaystyle{ \alpha_p(V_m) = p_\infty(V_m)/\tau_p }[/math]
- [math]\displaystyle{ \beta_p(V_m) = (1 - p_\infty(V_m))/\tau_p. }[/math]
[math]\displaystyle{ p_\infty }[/math] and [math]\displaystyle{ (1-p_\infty) }[/math] are the steady state values for activation and inactivation, respectively, and are usually represented by Boltzmann equations as functions of [math]\displaystyle{ V_m }[/math]. In the original paper by Hodgkin and Huxley,[1] the functions [math]\displaystyle{ \alpha }[/math] and [math]\displaystyle{ \beta }[/math] are given by
- [math]\displaystyle{ \begin{array}{lll} \alpha_n(V_m) = \frac{0.01(10-V)}{\exp\big(\frac{10-V}{10}\big)-1} & \alpha_m(V_m) = \frac{0.1(25-V)}{\exp\big(\frac{25-V}{10}\big)-1} & \alpha_h(V_m) = 0.07\exp\bigg(-\frac{V}{20}\bigg)\\ \beta_n(V_m) = 0.125\exp\bigg(-\frac{V}{80}\bigg) & \beta_m(V_m) = 4\exp\bigg(-\frac{V}{18}\bigg) & \beta_h(V_m) = \frac{1}{\exp\big(\frac{30-V}{10}\big) + 1} \end{array} }[/math]
where [math]\displaystyle{ V = V_{rest} - V_m }[/math] denotes the negative depolarization in mV.
[math]\displaystyle{ p_\infty }[/math] 和 [math]\displaystyle{ (1-p_\infty) }[/math]分别是激活和失活的稳态值,通常用玻尔兹曼方程表示为[math]\displaystyle{ V_m }[/math]的函数。
在 Hodgkin 和 Huxley 的原始论文[1]中,[math]\displaystyle{ \alpha }[/math]和[math]\displaystyle{ \beta }[/math]函数如下给出
[math]\displaystyle{ \begin{array}{lll}
\alpha_n(V_m) = \frac{0.01(10-V)}{\exp\big(\frac{10-V}{10}\big)-1} & \alpha_m(V_m) = \frac{0.1(25-V)}{\exp\big(\frac{25-V}{10}\big)-1} & \alpha_h(V_m) = 0.07\exp\bigg(-\frac{V}{20}\bigg)\\
\beta_n(V_m) = 0.125\exp\bigg(-\frac{V}{80}\bigg) & \beta_m(V_m) = 4\exp\bigg(-\frac{V}{18}\bigg) & \beta_h(V_m) = \frac{1}{\exp\big(\frac{30-V}{10}\big) + 1}
\end{array} }[/math]
其中[math]\displaystyle{ V = V_{rest} - V_m }[/math] 表示负去极化,单位为mV。
While in many current software programs,[2] Hodgkin–Huxley type models generalize [math]\displaystyle{ \alpha }[/math] and [math]\displaystyle{ \beta }[/math] to
在当前的软件程序中[3],霍奇金-赫胥黎类模型将[math]\displaystyle{ \alpha }[/math]和>[math]\displaystyle{ \beta }[/math]归纳为
- [math]\displaystyle{ \frac{A_p(V_m-B_p)}{\exp\big(\frac{V_m-B_p}{C_p}\big)-D_p} }[/math]
In order to characterize voltage-gated channels, the equations are fit to voltage clamp data. For a derivation of the Hodgkin–Huxley equations under voltage-clamp, see.[4] Briefly, when the membrane potential is held at a constant value (i.e., voltage-clamp), for each value of the membrane potential the nonlinear gating equations reduce to equations of the form:
为了刻画电压门控通道,该方程拟合自电压钳数据。关于电压钳下Hodgkin-Huxley方程的推导,请参阅[4]。简单来说,当膜电位保持为一个恒定值(即电压钳取值)时,对于膜电位的每个值,非线性门控方程可以归结为以下形式的方程:
- [math]\displaystyle{ m(t) = m_{0} - [ (m_{0}-m_{\infty})(1 - e^{-t/\tau_m})]\, }[/math]
- [math]\displaystyle{ h(t) = h_{0} - [ (h_{0}-h_{\infty})(1 - e^{-t/\tau_h})]\, }[/math]
- [math]\displaystyle{ n(t) = n_{0} - [ (n_{0}-n_{\infty})(1 - e^{-t/\tau_n})]\, }[/math]
Thus, for every value of membrane potential [math]\displaystyle{ V_{m} }[/math] the sodium and potassium currents can be described by
因此,对于膜电位的每个值[math]\displaystyle{ V_m }[/math],钠电流和钾电流可以描述为
- [math]\displaystyle{ I_\mathrm{Na}(t)=\bar{g}_\mathrm{Na} m(V_m)^3h(V_m)(V_m-E_\mathrm{Na}), }[/math]
- [math]\displaystyle{ I_\mathrm{K}(t)=\bar{g}_\mathrm{K} n(V_m)^4(V_m-E_\mathrm{K}). }[/math]
In order to arrive at the complete solution for a propagated action potential, one must write the current term I on the left-hand side of the first differential equation in terms of V, so that the equation becomes an equation for voltage alone. The relation between I and V can be derived from cable theory and is given by
为了得到传导的动作电位的完全解, 必须将第一个微分方程左侧的电流项 [math]\displaystyle{ I }[/math]写为[math]\displaystyle{ V }[/math]的形式,使方程成为单独的电压方程。[math]\displaystyle{ I }[/math]和[math]\displaystyle{ V }[/math]之间的关系可以从电缆理论中推导出来,即
- [math]\displaystyle{ I = \frac{a}{2R}\frac{\partial^2V}{\partial x^2}, }[/math]
where a is the radius of the axon, R is the specific resistance of the axoplasm, and x is the position along the nerve fiber. Substitution of this expression for I transforms the original set of equations into a set of partial differential equations, because the voltage becomes a function of both x and t.
其中[math]\displaystyle{ a }[/math]是轴突的半径,[math]\displaystyle{ R }[/math]是轴浆的比电阻,[math]\displaystyle{ x }[/math]是沿神经纤维的位置。用这个表达式代替[math]\displaystyle{ I }[/math],将原来的方程组转变为一组偏微分方程,因为电压变为[math]\displaystyle{ x }[/math]和[math]\displaystyle{ t }[/math]的函数。
The Levenberg–Marquardt algorithm is often used to fit these equations to voltage-clamp data.[5]
通常采用 Levenberg-Marquardt 算法通过电压钳数据拟合这些方程。
While the original experiments treated only sodium and potassium channels, the Hodgkin–Huxley model can also be extended to account for other species of ion channels.
虽然最初的实验只处理钠和钾通道,Hodgkin-Huxley模型也可以扩展到其他种类的离子通道。
漏通道
Leak channels account for the natural permeability of the membrane to ions and take the form of the equation for voltage-gated channels, where the conductance [math]\displaystyle{ g_{leak} }[/math] is a constant. Thus, the leak current due to passive leak ion channels in the Hodgkin-Huxley formalism is [math]\displaystyle{ I_l=g_{leak}(V-V_{leak}) }[/math].
漏通道解释了膜对离子的天然渗透性,其形式为电压门控通道方程,其中电导[math]\displaystyle{ g_{leak} }[/math]为常数。因此,在Hodgkin-Huxley公式中,被动漏离子通道引起的漏电流为[math]\displaystyle{ I_l=g_{leak}(V-V_{leak}) }[/math]。
泵和交换器
The membrane potential depends upon the maintenance of ionic concentration gradients across it. The maintenance of these concentration gradients requires active transport of ionic species. The sodium-potassium and sodium-calcium exchangers are the best known of these. Some of the basic properties of the Na/Ca exchanger have already been well-established: the stoichiometry of exchange is 3 Na+: 1 Ca2+ and the exchanger is electrogenic and voltage-sensitive. The Na/K exchanger has also been described in detail, with a 3 Na+: 2 K+ stoichiometry.[6][7]
膜电位取决于其跨膜离子浓度梯度的保持。维持这些浓度梯度需要这几种离子的主动运输。其中钠钾交换器和钠钙交换器最为著名。钠钙交换器的一些基本性质已得到公认: 交换的化学计量比为 3 Na+: 1 Ca2+,且具有生电性和电压敏感性。文献中还详细描述了Na/K交换器,它具有 3 Na+: 2 K+的化学计量比。[6][7]
数学性质
The Hodgkin–Huxley model can be thought of as a differential equation system with four state variables, [math]\displaystyle{ V_m(t), n(t), m(t) }[/math], and [math]\displaystyle{ h(t) }[/math], that change with respect to time [math]\displaystyle{ t }[/math]. The system is difficult to study because it is a nonlinear system and cannot be solved analytically. However, there are many numerical methods available to analyze the system. Certain properties and general behaviors, such as limit cycles, can be proven to exist.
可以认为Hodgkin-Huxley 模型是一个具有4个状态变量[math]\displaystyle{ V_m(t),n(t),m(t) }[/math],和[math]\displaystyle{ h(t) }[/math]的微分方程系统,它们随着时间[math]\displaystyle{ t }[/math]变化。这个系统很难研究,因为它是一个非线性系统,无法用解析法求解。然而,可以用许多数值方法分析该系统。可以证明某些性质和一般行为(如极限环)是存在的。
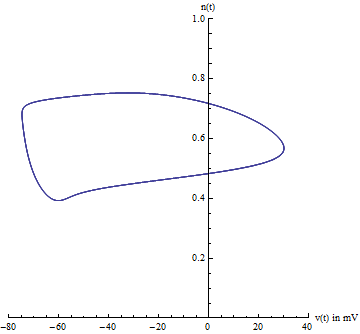
{kind=link}
中心流形
Because there are four state variables, visualizing the path in phase space can be difficult. Usually two variables are chosen, voltage [math]\displaystyle{ V_m(t) }[/math] and the potassium gating variable [math]\displaystyle{ n(t) }[/math], allowing one to visualize the limit cycle. However, one must be careful because this is an ad-hoc method of visualizing the 4-dimensional system. This does not prove the existence of the limit cycle.
由于有四个状态变量,想象相空间中的路径会比较困难。通常选择两个变量,电压[math]\displaystyle{ V_m(t) }[/math]和钾门控变量[math]\displaystyle{ n(t) }[/math],这样就能想象出极限环。但是,要注意这只是一个想象四维系统的特殊方法,并不能证明极限环的存在性。
A better projection can be constructed from a careful analysis of the Jacobian of the system, evaluated at the equilibrium point. Specifically, the eigenvalues of the Jacobian are indicative of the center manifold's existence. Likewise, the eigenvectors of the Jacobian reveal the center manifold's orientation. The Hodgkin–Huxley model has two negative eigenvalues and two complex eigenvalues with slightly positive real parts. The eigenvectors associated with the two negative eigenvalues will reduce to zero as time t increases. The remaining two complex eigenvectors define the center manifold. In other words, the 4-dimensional system collapses onto a 2-dimensional plane. Any solution starting off the center manifold will decay towards the center manifold. Furthermore, the limit cycle is contained on the center manifold.
在系统的平衡点处对雅可比矩阵仔细分析,可以构造出一个更好的投影。 具体来说,雅可比矩阵的特征值指示中心流形的存在。 同样,雅可比的特征向量揭示了中心流形的方向。 Hodgkin-Huxley 模型有两个负的特征值和两个具有轻微取正的实部的复特征值。 随着时间 t 的增加,与两个负特征值相关的特征向量将减少到零。 剩下的两个复特征向量定义了中心流形。换句话说,这个四维系统会坍缩到一个二维平面上。 任何开始于中心流形的解将衰减至中心流形。 此外,极限环包含在中心流形上。
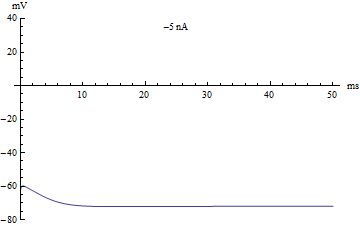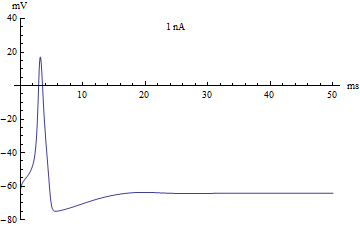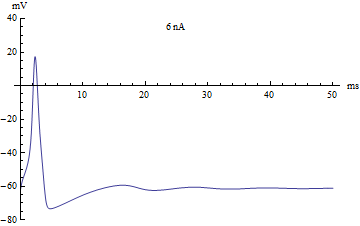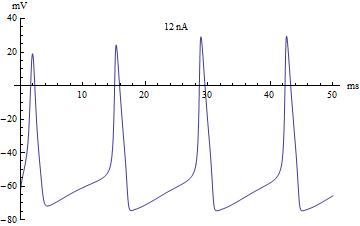
{kind=link}
分岔
If the injected current [math]\displaystyle{ I }[/math] were used as a bifurcation parameter, then the Hodgkin–Huxley model undergoes a Hopf bifurcation. As with most neuronal models, increasing the injected current will increase the firing rate of the neuron. One consequence of the Hopf bifurcation is that there is a minimum firing rate. This means that either the neuron is not firing at all (corresponding to zero frequency), or firing at the minimum firing rate. Because of the all-or-none principle, there is no smooth increase in action potential amplitude, but rather there is a sudden "jump" in amplitude. The resulting transition is known as a canard.
如果将注入电流 [math]\displaystyle{ I }[/math] 作为分岔参数,那么 Hodgkin-Huxley 模型将经历一个霍普夫分岔。和大多数神经元模型一样,增加注入电流会增加神经元的发放率。霍普夫分岔的一个结果就是存在一个最低的发放率。这意味着神经元要么根本没有发放(对应于零频率) ,要么以最低发放率放电。由于全或无原理,动作电位的幅度不存在平稳的增加,而是幅度上的突然“跳跃”。由此产生的转变被称为鸭解。
改进与可替代模型
The Hodgkin–Huxley model is regarded as one of the great achievements of 20th-century biophysics. Nevertheless, modern Hodgkin–Huxley-type models have been extended in several important ways:
- Additional ion channel populations have been incorporated based on experimental data.
- The Hodgkin–Huxley model has been modified to incorporate transition state theory and produce thermodynamic Hodgkin–Huxley models.[8]
- Models often incorporate highly complex geometries of dendrites and axons, often based on microscopy data.
- Stochastic models of ion-channel behavior, leading to stochastic hybrid systems.[9]
- The Poisson–Nernst–Planck (PNP) model is based on a mean-field approximation of ion interactions and continuum descriptions of concentration and electrostatic potential.[10]
霍奇金-赫胥黎模型被认为是20世纪生物物理学的伟大成就之一。尽管如此,现代 Hodgkin-Huxley 型模型已经在几个重要方面得到了扩展:
- 根据实验数据引入了额外的离子通道群。
- 对Hodgkin-Huxley 模型加以修正,加入了过渡态理论,并产生了热力学 Hodgkin-Huxley 模型。[11]
- 通常基于显微镜数据,模型通常包含树突和轴突的高度复杂的几何形状。
- 离子通道行为的随机模型,导致随机混合系统。[9]
- Poisson-Nernst-Planck 模型是基于离子相互作用的平均场近似以及浓度和静电势的连续描述建立的。[12]
Several simplified neuronal models have also been developed (such as the FitzHugh–Nagumo model), facilitating efficient large-scale simulation of groups of neurons, as well as mathematical insight into dynamics of action potential generation. 一些简化的神经元模型(如 FitzHugh-Nagumo 模型)也发展了出来,它们有助于对神经元群进行高效的大规模模拟,以及对动作电位产生的动力学的数学洞察。
参见
参考文献
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 Hodgkin AL, Huxley AF (August 1952). "A quantitative description of membrane current and its application to conduction and excitation in nerve". The Journal of Physiology. 117 (4): 500–44. doi:10.1113/jphysiol.1952.sp004764. PMC 1392413. PMID 12991237.
- ↑ Nelson ME (2005) Electrophysiological Models In: Databasing the Brain: From Data to Knowledge. (S. Koslow and S. Subramaniam, eds.) Wiley, New York, pp. 285–301
- ↑ Nelson ME (2005) Electrophysiological Models In: Databasing the Brain: From Data to Knowledge. (S. Koslow and S. Subramaniam, eds.) Wiley, New York, pp. 285–301
- ↑ 4.0 4.1 Gray, Daniel Johnston; Wu, Samuel Miao-Sin (1997). Foundations of cellular neurophysiology (3rd ed.). Cambridge, Massachusetts [u.a.]: MIT Press. ISBN 978-0-262-10053-3.
- ↑ Krapivin, Vladimir F.; Varotsos, Costas A.; Soldatov, Vladimir Yu. (2015). New Ecoinformatics Tools in Environmental Science : Applications and Decision-making. Springer. pp. 37–38. ISBN 9783319139784. https://www.google.com/books/edition/New_Ecoinformatics_Tools_in_Environmenta/bWpnBgAAQBAJ?hl=en&gbpv=1&pg=PA37.
- ↑ 6.0 6.1 Rakowski RF, Gadsby DC, De Weer P (May 1989). "Stoichiometry and voltage dependence of the sodium pump in voltage-clamped, internally dialyzed squid giant axon". The Journal of General Physiology. 93 (5): 903–41. doi:10.1085/jgp.93.5.903. PMC 2216238. PMID 2544655.
- ↑ 7.0 7.1 Hille, Bertil (2001). Ion channels of excitable membranes (3rd ed.). Sunderland, Massachusetts: Sinauer. ISBN 978-0-87893-321-1.
- ↑ Forrest, M. D. (May 2014). "Can the Thermodynamic Hodgkin–Huxley Model of Voltage-Dependent Conductance Extrapolate for Temperature?" (PDF). Computation. 2 (2): 47–60. doi:10.3390/computation2020047.
- ↑ 9.0 9.1 Pakdaman, K.; Thieullen, M.; Wainrib, G. (2010). "Fluid limit theorems for stochastic hybrid systems with applications to neuron models". Adv. Appl. Probab. 42 (3): 761–794. arXiv:1001.2474. Bibcode:2010arXiv1001.2474P. doi:10.1239/aap/1282924062. S2CID 18894661.
- ↑ Zheng, Q.; Wei, G. W. (May 2011). "Poisson-Boltzmann-Nernst-Planck model". Journal of Chemical Physics. 134 (19): 194101. Bibcode:2011JChPh.134s4101Z. doi:10.1063/1.3581031. PMC 3122111. PMID 21599038.
- ↑ Forrest, M. D. (May 2014). "Can the Thermodynamic Hodgkin–Huxley Model of Voltage-Dependent Conductance Extrapolate for Temperature?" (PDF). Computation. 2 (2): 47–60. doi:10.3390/computation2020047.
- ↑ Zheng, Q.; Wei, G. W. (May 2011). "Poisson-Boltzmann-Nernst-Planck model". Journal of Chemical Physics. 134 (19): 194101. Bibcode:2011JChPh.134s4101Z. doi:10.1063/1.3581031. PMC 3122111. PMID 21599038.
进一步阅读
- Hodgkin AL, Huxley AF (April 1952). "Currents carried by sodium and potassium ions through the membrane of the giant axon of Loligo". The Journal of Physiology. 116 (4): 449–72. doi:10.1113/jphysiol.1952.sp004717. PMC 1392213. PMID 14946713.
- Hodgkin AL, Huxley AF (April 1952). "The components of membrane conductance in the giant axon of Loligo". The Journal of Physiology. 116 (4): 473–96. doi:10.1113/jphysiol.1952.sp004718. PMC 1392209. PMID 14946714.
- Hodgkin AL, Huxley AF (April 1952). "The dual effect of membrane potential on sodium conductance in the giant axon of Loligo". The Journal of Physiology. 116 (4): 497–506. doi:10.1113/jphysiol.1952.sp004719. PMC 1392212. PMID 14946715.
- Hodgkin AL, Huxley AF (August 1952). "A quantitative description of membrane current and its application to conduction and excitation in nerve". The Journal of Physiology. 117 (4): 500–44. doi:10.1113/jphysiol.1952.sp004764. PMC 1392413. PMID 12991237.
- Hodgkin AL, Huxley AF, Katz B (April 1952). "Measurement of current-voltage relations in the membrane of the giant axon of Loligo". The Journal of Physiology. 116 (4): 424–48. doi:10.1113/jphysiol.1952.sp004716. PMC 1392219. PMID 14946712.
外部链接
- Interactive Javascript simulation of the HH model Runs in any HTML5 – capable browser. Allows for changing the parameters of the model and current injection.
- Interactive Java applet of the HH model Parameters of the model can be changed as well as excitation parameters and phase space plottings of all the variables is possible.
- Direct link to Hodgkin–Huxley model and a Description in BioModels Database
- Neural Impulses: The Action Potential In Action by Garrett Neske, The Wolfram Demonstrations Project
- Interactive Hodgkin–Huxley model by Shimon Marom, The Wolfram Demonstrations Project
- ModelDB A computational neuroscience source code database containing 4 versions (in different simulators) of the original Hodgkin–Huxley model and hundreds of models that apply the Hodgkin–Huxley model to other channels in many electrically excitable cell types.
- Several articles about the stochastic version of the model and its link with the original one.
- Interactive Javascript simulation of the HH model Runs in any HTML5 – capable browser. Allows for changing the parameters of the model and current injection.
- Interactive Java applet of the HH model Parameters of the model can be changed as well as excitation parameters and phase space plottings of all the variables is possible.
- Direct link to Hodgkin–Huxley model and a Description in BioModels Database
- Neural Impulses: The Action Potential In Action by Garrett Neske, The Wolfram Demonstrations Project
- Interactive Hodgkin–Huxley model by Shimon Marom, The Wolfram Demonstrations Project
- ModelDB A computational neuroscience source code database containing 4 versions (in different simulators) of the original Hodgkin–Huxley model and hundreds of models that apply the Hodgkin–Huxley model to other channels in many electrically excitable cell types.
- Several articles about the stochastic version of the model and its link with the original one.
编者推荐
《神经科学的数学原理》
Category:Nonlinear systems 非线性系统
Category:Electrophysiology 电生理学
Category:Ion channels 离子通道
Category:Computational neuroscience 计算神经科学
类别: 非线性系统类别: 电生理学类别: 离子通道类别: 计算神经科学
This page was moved from wikipedia:en:Hodgkin–Huxley model. Its edit history can be viewed at 霍奇金-赫胥黎模型/edithistory