树突
树突(源于希腊语 δνδρν déndron,树的意思),是神经细胞的分枝状的原生质延伸,对其他神经细胞传递的电化学刺激传播到该细胞的胞体。电刺激信号由上游神经元(通常通过其轴突)经树突分支各处的突触传递到树突上。树突在整合这些突触输入以及决定神经元动作电位发生起着关键作用[1]。树突分枝形成,是个多步骤的生物过程,藉此神经元形成新的树突分支和分叉,以创建新突触[1]。树突的形态如分支密度和分组模式与神经元的功能密切相关。树突畸形也与神经系统功能损伤有很高关联。与树突畸形有关的疾病包括自闭症、抑郁症、精神分裂症、唐氏综合症和焦虑症。
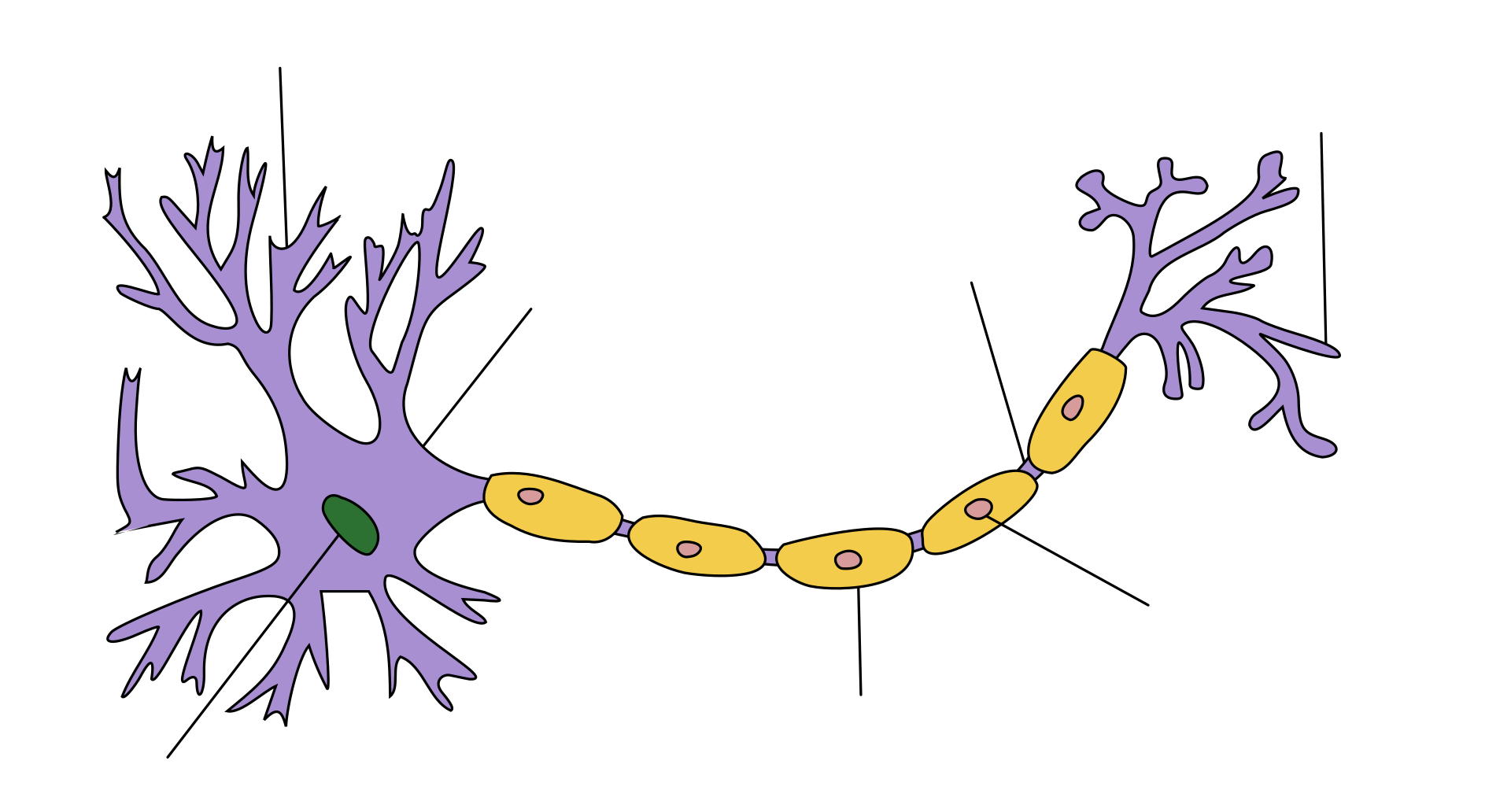
Certain classes of dendrites contain small projections referred to as dendritic spines that increase receptive properties of dendrites to isolate signal specificity. Increased neural activity and the establishment of long-term potentiation at dendritic spines change the sizes, shape, and conduction. This ability for dendritic growth is thought to play a role in learning and memory formation. There can be as many as 15,000 spines per cell, each of which serves as a postsynaptic process for individual presynaptic axons. Dendritic branching can be extensive and in some cases is sufficient to receive as many as 100,000 inputs to a single neuron. Dendrites are one of two types of protoplasmic protrusions that extrude from the cell body of a neuron, the other type being an axon. Axons can be distinguished from dendrites by several features including shape, length, and function. Dendrites often taper off in shape and are shorter, while axons tend to maintain a constant radius and be relatively long. Typically, axons transmit electrochemical signals and dendrites receive the electrochemical signals, although some types of neurons in certain species lack axons and simply transmit signals via their dendrites. Dendrites provide an enlarged surface area to receive signals from the terminal buttons of other axons, and the axon also commonly divides at its far end into many branches (telodendria) each of which ends in a nerve terminal, allowing a chemical signal to pass simultaneously to many target cells.
Typically, when an electrochemical signal stimulates a neuron, it occurs at a dendrite and causes changes in the electrical potential across the neuron's plasma membrane. This change in the membrane potential will passively spread across the dendrite but becomes weaker with distance without an action potential. An action potential propagates the electrical activity along the membrane of the neuron's dendrites to the cell body and then afferently down the length of the axon to the axon terminal, where it triggers the release of neurotransmitters into the synaptic cleft. However, synapses involving dendrites can also be axodendritic, involving an axon signaling to a dendrite, or dendrodendritic, involving signaling between dendrites. An autapse is a synapse in which the axon of one neuron transmits signals to its own dendrites.
某些类型的树突具有称为树突棘的小突起,增加树突的信号接收特性,通过隔离增强信号的特异性。树突棘的神经活动增强和长时程增强会改变其大小、形状和传导性能。这种树突生长能力被认为在学习和记忆形成中起着重要作用。每个细胞可以有多达 15,000 个树突棘,每个都可以作为单个突触前轴突的突触后突起[2]。树突分支可以非常庞大,足以为单个神经元接收多达 100,000 个输入[3]。树突是从神经元细胞体延伸出来的两种原生质突起之一,另一种是轴突。轴突与树突可以通过几个特征来区分,包括形状、长度和功能。树突有逐渐变细的形状,更短,而轴突倾向于保持恒定的半径,相对较长。通常,轴突传递电化学信号,树突接收电化学信号,尽管某些种类的神经元没有轴突,仅仅通过树突传递信号[4] 。树突提供了扩大的表面区域来接收来自其他轴突末端的信号,轴突也通常在其远端形成许多分支(终树突),每个分支形成一个神经末梢,允许一种化学信号同时传递给许多目标细胞[3] 。当电化学信号,通常在树突部位,刺激神经元,并引起神经元质膜电位的变化。这种膜电位的变化会在整个树突进行被动扩布,但随着距离的增加而变弱,不会产生动作电位。动作电位沿着神经元树突的细胞膜传递电活动到胞体,然后沿着轴突传递到轴突末梢,在那里触发神经递质的释放到突触间隙[3]。然而,包含树突的突触也可以是轴树突触,即从轴突向树突传递信号,也可以是树树突触,即树突之间传递信号[5]。自突触是一种神经元的轴突将信号传递给自身树突的突触。
There are three main types of neurons; multipolar, bipolar, and unipolar. Multipolar neurons, such as the one shown in the image, are composed of one axon and many dendritic trees. Pyramidal cells are multipolar cortical neurons with pyramid shaped cell bodies and large dendrites called apical dendrites that extend to the surface of the cortex. Bipolar neurons have one axon and one dendritic tree at opposing ends of the cell body. Unipolar neurons have a stalk that extends from the cell body that separates into two branches with one containing the dendrites and the other with the terminal buttons. Unipolar dendrites are used to detect sensory stimuli such as touch or temperature.[5][6][7]
There are three main types of neurons; multipolar, bipolar, and unipolar. Multipolar neurons, such as the one shown in the image, are composed of one axon and many dendritic trees. Pyramidal cells are multipolar cortical neurons with pyramid shaped cell bodies and large dendrites called apical dendrites that extend to the surface of the cortex. Bipolar neurons have one axon and one dendritic tree at opposing ends of the cell body. Unipolar neurons have a stalk that extends from the cell body that separates into two branches with one containing the dendrites and the other with the terminal buttons. Unipolar dendrites are used to detect sensory stimuli such as touch or temperature.
神经元有三种主要类型:多极、双极和单极。多极神经元,如图所示,由一个轴突和许多树突组成。锥体细胞是多极皮层神经元,具有锥形的胞体和延伸到皮层表面的称为顶树突的大树突。双极神经元在胞体两端分别有一个轴突和一个树突树。单极神经元有一根柄从细胞体延伸出来,分成两个分支,一个分支包含树突,另一个分支包含末梢。单极树突被用来检测感觉刺激,如触摸或温度。
历史
The term dendrites was first used in 1889 by Wilhelm His to describe the number of smaller "protoplasmic processes" that were attached to a nerve cell.[8] German anatomist Otto Friedrich Karl Deiters is generally credited with the discovery of the axon by distinguishing it from the dendrites.
The term dendrites was first used in 1889 by Wilhelm His to describe the number of smaller "protoplasmic processes" that were attached to a nerve cell. German anatomist Otto Friedrich Karl Deiters is generally credited with the discovery of the axon by distinguishing it from the dendrites.
树突(dendrite)这一术语最早是在 1889 年被 Wilhelm His 用来描述神经细胞相连的较小的“原生质突起”的数量。德国解剖学家 Otto Friedrich Karl Deiters 被认为发现了轴突,是他首先将轴突与树突区分开。
Some of the first intracellular recordings in a nervous system were made in the late 1930s by Kenneth S. Cole and Howard J. Curtis. Swiss Rüdolf Albert von Kölliker and German Robert Remak were the first to identify and characterize the axonal initial segment. Alan Hodgkin and Andrew Huxley also employed the squid giant axon (1939) and by 1952 they had obtained a full quantitative description of the ionic basis of the action potential, leading the formulation of the Hodgkin–Huxley model. Hodgkin and Huxley were awarded jointly the Nobel Prize for this work in 1963. The formulas detailing axonal conductance were extended to vertebrates in the Frankenhaeuser–Huxley equations. Louis-Antoine Ranvier was the first to describe the gaps or nodes found on axons and for this contribution these axonal features are now commonly referred to as the Nodes of Ranvier. Santiago Ramón y Cajal, a Spanish anatomist, proposed that axons were the output components of neurons. He also proposed that neurons were discrete cells that communicated with each other via specialized junctions, or spaces, between cells, now known as a synapse. Ramón y Cajal improved a silver staining process known as Golgi's method, which had been developed by his rival, Camillo Golgi.
1930 年代末,Kenneth S. Cole 和 Howard J. Curtis 开展了最早的神经系统胞内电生理记录。瑞士人 Rüdolf Albert von Kölliker 和德国人 German Robert Remak 首先对轴突起始段进行了鉴定和性质研究。1952 年,Alan Hodgkin 和 Andrew Huxley 也使用了乌贼巨突触(1939年),到 1952 年他们得到动作电位的离子机制的完整定量描述,从而建立了 Hodgkin-Huxley 模型。Hodgkin 和 Huxley 因这项工作在 1963 年共同获得诺贝尔奖。Frankenhaeuser-Huxley 方程式则将详细描述轴突传导的公式推广到脊椎动物。Louis-Antoine Ranvier 首次描述了轴突上的不连续或饥结,因此这些轴突特征现在通常被称为郎飞结。西班牙解剖学家 Ramón y Cajal 提出轴突是神经元输出的部分[9]。他还提出,神经元是离散的细胞,通过细胞间的特殊连接或空隙彼此沟通,今天称为突触的结构进行相互通信。Ramón y Cajal 改进了他的竞争对手 Camillo Golgi 发明的高尔基染色法[10]。
Dendrite development 树突发育

{kind=link}
{kind=link}
During the development of dendrites, several factors can influence differentiation. These include modulation of sensory input, environmental pollutants, body temperature, and drug use.[11] For example, rats raised in dark environments were found to have a reduced number of spines in pyramidal cells located in the primary visual cortex and a marked change in distribution of dendrite branching in layer 4 stellate cells.[12] Experiments done in vitro and in vivo have shown that the presence of afferents and input activity per se can modulate the patterns in which dendrites differentiate.[13]
在树突的发育过程中,有几个因素可以影响这种分化,包括感觉输入调控、环境污染物、体温和药物使用[11] 。例如,在黑暗环境中长大的大鼠,初级视皮层的锥体细胞的树突棘数量减少,在第 4 层星状细胞中树突分支的分布有明显的不同[12] 。体外和体内实验表明,传入神经和传入神经活动本身可以调节树突分化的模式[13]。
Little is known about the process by which dendrites orient themselves in vivo and are compelled to create the intricate branching pattern unique to each specific neuronal class. One theory on the mechanism of dendritic arbor development is the Synaptotropic Hypothesis. The synaptotropic hypothesis proposes that input from a presynaptic to a postsynaptic cell (and maturation of excitatory synaptic inputs) eventually can change the course of synapse formation at dendritic and axonal arbors.[14] This synapse formation is required for the development of neuronal structure in the functioning brain. A balance between metabolic costs of dendritic elaboration and the need to cover receptive field presumably determine the size and shape of dendrites. A complex array of extracellular and intracellular cues modulates dendrite development including transcription factors, receptor-ligand interactions, various signaling pathways, local translational machinery, cytoskeletal elements, Golgi outposts and endosomes. These contribute to the organization of the dendrites on individual cell bodies and the placement of these dendrites in the neuronal circuitry. For example, it was shown that β-actin zipcode binding protein 1 (ZBP1) contributes to proper dendritic branching. Other important transcription factors involved in the morphology of dendrites include CUT, Abrupt, Collier, Spineless, ACJ6/drifter, CREST, NEUROD1, CREB, NEUROG2 etc. Secreted proteins and cell surface receptors includes neurotrophins and tyrosine kinase receptors, BMP7, Wnt/dishevelled, EPHB 1–3, Semaphorin/plexin-neuropilin, slit-robo, netrin-frazzled, reelin. Rac, CDC42 and RhoA serve as cytoskeletal regulators and the motor protein includes KIF5, dynein, LIS1. Important secretory and endocytic pathways controlling the dendritic development include DAR3 /SAR1, DAR2/Sec23, DAR6/Rab1 etc. All these molecules interplay with each other in controlling dendritic morphogenesis including the acquisition of type specific dendritic arborization, the regulation of dendrite size and the organization of dendrites emanating from different neurons.[1][15]
树突如何在体内定向,并产生特定神经元类别特有的复杂分支模式,这个过程我们知之甚少。树突分支发育机制的一个理论是突触营养假设。突触营养假设认为,突触前细胞向突触后细胞的输入(以及兴奋性突触输入的成熟)最终可以改变树突和轴突末梢的分支形成突触的进程[14] 。这种突触形成对工作中的大脑的神经元结构发育所必需的。树突复杂分支的形成的代谢成本和覆盖感受野的需要之间的平衡大体决定了树突的大小和形状。一系列复杂的胞外和胞内信号,包括转录因子、受体-配体相互作用、各种信号通路、局部翻译机制、细胞骨架元件、高尔基前哨和内涵体,都会调节树突发育。这些促进了单个细胞胞体上树突的组织以及这些树突在神经回路中的定位。例如,研究表明 β 肌动蛋白结合蛋白1(β-actin zipcode binding protein 1,ZBP1)在树突形成正常分支中有作用。其他与树突形态有关的重要转录因子包括 CUT、stump、Collier、Spineless、ACJ6/drifter、CREST、NEUROD1、CREB、neurog2 等。分泌蛋白和细胞表面受体包括神经营养因子和酪氨酸受体激酶、BMP7、Wnt/dishevelled、EPHB 1-3、 Semaphorin/plexin-neuropilin、slit-robo、netrin-frazzled、reelin。Rac, CDC42 和 RhoA 作为细胞骨架调节分子,马达蛋白包括 KIF5、dynein、LIS1。控制树突发育的重要分泌和内吞途径包括 DAR3/SAR1、DAR2/Sec23、DAR6/rab1 等。所有这些分子在控制树突形态发生中相互作用,包括细胞类型特异的树突分支、树突大小和不同神经元树突的组织[1][15]。
Electrical properties 电性质
The structure and branching of a neuron's dendrites, as well as the availability and variation of voltage-gated ion conductance, strongly influences how the neuron integrates the input from other neurons. This integration is both temporal, involving the summation of stimuli that arrive in rapid succession, as well as spatial, entailing the aggregation of excitatory and inhibitory inputs from separate branches.[16]
The structure and branching of a neuron's dendrites, as well as the availability and variation of voltage-gated ion conductance, strongly influences how the neuron integrates the input from other neurons. This integration is both temporal, involving the summation of stimuli that arrive in rapid succession, as well as spatial, entailing the aggregation of excitatory and inhibitory inputs from separate branches.
神经元树突的结构和分支模式,以及电压门控离子通道的表达与种类,强烈地影响着神经元如何整合来自其他神经元的输入。这种整合既包含时间整合,即快速连续到达的刺激的总和,也包含空间整合,即来自不同分支的兴奋性和抑制性输入的聚合[16]。
Dendrites were once thought to merely convey electrical stimulation passively. This passive transmission means that voltage changes measured at the cell body are the result of activation of distal synapses propagating the electric signal towards the cell body without the aid of voltage-gated ion channels. Passive cable theory describes how voltage changes at a particular location on a dendrite transmit this electrical signal through a system of converging dendrite segments of different diameters, lengths, and electrical properties. Based on passive cable theory one can track how changes in a neuron's dendritic morphology impacts the membrane voltage at the cell body, and thus how variation in dendrite architectures affects the overall output characteristics of the neuron.[17][18]
树突之前被认为只是被动地传导电刺激。这种被动传输意味着,在细胞体上测量到的电压变化是远端突触激活产生的电信号,在没有电压门控离子通道的帮助下,向胞体传播的结果。无源电缆理论描述了树突特定位置的电压变化如何通过不同直径、长度和电性质的汇聚的树突系统传输这种电信号。基于被动电缆理论,人们可以探究神经元树突形态的变化如何影响胞体的膜电位,从而了解树突的结构体系变化如何影响神经元的整体输出特性[17][18]。
Electrochemical signals are propagated by action potentials that utilize intermembrane voltage-gated ion channels to transport sodium ions, calcium ions, and potassium ions. Each ion species has its own corresponding protein channel located in the lipid bilayer of the cell membrane. The cell membrane of neurons covers the axons, cell body, dendrites, etc. The protein channels can differ between chemical species in the amount of required activation voltage and the activation duration.[3]
电化学信号可以利用动作电位进行传播,动作电位使用电压门控离子通道跨膜转运钠离子、钙离子和钾离子。每种离子在细胞膜的脂质双分子层中都有自己对应的蛋白通道。神经元的细胞膜覆盖轴突、胞体、树突等。不同离子的蛋白质通道具有不同的激活电位和持续时间[3]。
Action potentials in animal cells are generated by either sodium-gated or calcium-gated ion channels in the plasma membrane. These channels are closed when the membrane potential is near to, or at, the resting potential of the cell. The channels will start to open if the membrane potential increases, allowing sodium or calcium ions to flow into the cell. As more ions enter the cell, the membrane potential continues to rise. The process continues until all of the ion channels are open, causing a rapid increase in the membrane potential that then triggers the decrease in the membrane potential. The depolarizing is caused by the closing of the ion channels that prevent sodium ions from entering the neuron, and they are then actively transported out of the cell. Potassium channels are then activated, and there is an outward flow of potassium ions, returning the electrochemical gradient to the resting potential. After an action potential has occurred, there is a transient negative shift, called the afterhyperpolarization or refractory period, due to additional potassium currents. This is the mechanism that prevents an action potential from traveling back the way it just came.[3][19]
动物细胞的动作电位是由质膜上的钠门控或钙门控的离子通道产生的。这些通道在膜电位接近细胞的静息电位时关闭。随着膜电位增加,通道将开始开放,使钠离子或钙离子流入细胞。随着更多的离子进入细胞,膜电位会继续上升。这个过程一直持续到所有的离子通道都打开,导致膜电位的快速增加,然后触发膜电位的减少。去极化是因为钠离子打开,然后钠离子外流。然后激活钾离子通道,钾离子外流,将电化学梯度回到静息电位。动作电位发生后,由于额外的钾电流,膜电位会有一个短暂的负移,称为后超极化或不应期。这种机制可以阻止动作电位逆向传播[3][19]。
Another important feature of dendrites, endowed by their active voltage gated conductance, is their ability to send action potentials back into the dendritic arbor. Known as back-propagating action potentials, these signals depolarize the dendritic arbor and provide a crucial component toward synapse modulation and long-term potentiation. Furthermore, a train of back-propagating action potentials artificially generated at the soma can induce a calcium action potential (a dendritic spike) at the dendritic initiation zone in certain types of neurons.[citation needed]
树突的另一个重要特征,由其活跃电压门控电导赋予的,是能够将动作电位传回突出分支。这些被称为反向传播动作电位的信号使树突轴去极化,是进行突触调制和长时程增强的重要组成。此外,在胞体人为产生一串反向传播动作电位可以在特定类型的神经元的树突起始区诱导钙动作电位(树突发放)。
Plasticity 可塑性
Dendrites themselves appear to be capable of plastic changes during the adult life of animals, including invertebrates. Neuronal dendrites have various compartments known as functional units that are able to compute incoming stimuli. These functional units are involved in processing input and are composed of the subdomains of dendrites such as spines, branches, or groupings of branches. Therefore, plasticity that leads to changes in the dendrite structure will affect communication and processing in the cell. During development, dendrite morphology is shaped by intrinsic programs within the cell's genome and extrinsic factors such as signals from other cells. But in adult life, extrinsic signals become more influential and cause more significant changes in dendrite structure compared to intrinsic signals during development. In females, the dendritic structure can change as a result of physiological conditions induced by hormones during periods such as pregnancy, lactation, and following the estrous cycle. This is particularly visible in pyramidal cells of the CA1 region of the hippocampus, where the density of dendrites can vary up to 30%.[13]
树突在动物包括无脊椎动物的成年生命阶段似乎能够发生可塑性变化。神经元树突有许多被称为功能单元的区域,它们能够对传入的刺激进行计算。这些功能单元参与处理的输入,由树突的子域,如树突棘、分支或分支组组成。因此,引起树突结构变化的可塑性将影响细胞的通信和处理。在发育过程中,树突形态是由细胞基因组编码的内在程序和(来自其他细胞的信号)外在因子塑造的。但在成年阶段,相比于发育阶段,外源信号相比于内源信号,对树突结构的影响更为显著。在女性,树突结构可以因激素诱导的生理状态如怀孕、哺乳、动情周期期间而改变。这在海马 CA1 区的锥体细胞中尤其明显,起树突密度的变化可达30% [13]。
Notes
- ↑ 1.0 1.1 1.2 1.3 Urbanska, M.; Blazejczyk, M.; Jaworski, J. (2008). "Molecular basis of dendritic arborization". Acta Neurobiologiae Experimentalis. 68 (2): 264–288. PMID 18511961.
- ↑ Koch, C.; Zador, A. (February 1993). "The Function of Dendritic Spines: Devices Subserving Biochemical Rather Than Electrical Compartmentalization". The Journal of Neuroscience. 13 (2): 413–422. doi:10.1523/JNEUROSCI.13-02-00413.1993. PMC 6576662. PMID 8426220.
- ↑ 3.0 3.1 3.2 3.3 3.4 3.5 3.6 Alberts, Bruce (2009). Essential Cell Biology (3rd ed.). New York: Garland Science. ISBN 978-0-8153-4129-1.
- ↑ Yau, K. W. (1976). "Receptive fields, geometry and conduction block of sensory neurones in the central nervous system of the leech". The Journal of Physiology. 263 (3): 513–38. doi:10.1113/jphysiol.1976.sp011643. PMC 1307715. PMID 1018277.
- ↑ 5.0 5.1 Carlson, Neil R. (2013). Physiology of Behavior (11th ed.). Boston: Pearson. ISBN 978-0-205-23939-9.
- ↑ Pinel, John P.J. (2011). Biopsychology (8th ed.). Boston: Allyn & Bacon. ISBN 978-0-205-83256-9.
- ↑ Jan, Y. N.; Jan, L. Y. (2010). "Branching out: Mechanisms of dendritic arborization". Nature Reviews Neuroscience. 11 (5): 316–328. doi:10.1038/nrn2836. PMC 3079328. PMID 20404840.
- ↑ Finger, Stanley (1994). Origins of neuroscience : a history of explorations into brain function. Oxford University Press. pp. 44. ISBN 9780195146943. OCLC 27151391. "The nerve cell with its uninterrupted processes was described by Otto Friedrich Karl Deiters (1834-1863) in a work that was completed by Max Schultze (1825-1874) in 1865, two years after Deiters died of typhoid fever. This work portrayed the cell body with a single chief "axis cylinder" and a number of smaller "protoplasmic processes" (see figure 3.19). The latter would become known as "dendrites", a term coined by Wilhelm His (1831-1904) in 1889."
- ↑ Debanne, D; Campanac, E; Bialowas, A; Carlier, E; Alcaraz, G (Apr 2011). "Axon physiology" (PDF). Physiological Reviews. 91 (2): 555–602. doi:10.1152/physrev.00048.2009. PMID 21527732.
- ↑ López-Muñoz, F (October 2006). "Neuron theory, the cornerstone of neuroscience, on the centenary of the Nobel Prize award to Santiago Ramón y Cajal". Brain Research Bulletin. 70 (4–6): 391–405. doi:10.1016/j.brainresbull.2006.07.010. PMID 17027775. S2CID 11273256.
- ↑ 11.0 11.1 McEwen, Bruce S. (2010). "Stress, sex, and neural adaptation to a changing environment: mechanisms of neuronal remodeling". Annals of the New York Academy of Sciences. 1204: 38–59. Bibcode:2010NYASA1204...38M. doi:10.1111/j.1749-6632.2010.05568.x. PMC 2946089. PMID 20840167.
- ↑ 12.0 12.1 Borges, S.; Berry, M. (15 July 1978). "The effects of dark rearing on the development of the visual cortex of the rat". The Journal of Comparative Neurology. 180 (2): 277–300. doi:10.1002/cne.901800207. PMID 659662. S2CID 42749947.
- ↑ 13.0 13.1 13.2 13.3 Tavosanis, G. (2012). "Dendritic structural plasticity". Developmental Neurobiology. 72 (1): 73–86. doi:10.1002/dneu.20951. PMID 21761575. S2CID 2055017.
- ↑ 14.0 14.1 Cline, H; Haas, K (Mar 15, 2008). "The regulation of dendritic arbor development and plasticity by glutamatergic synaptic input: a review of the synaptotrophic hypothesis". The Journal of Physiology. 586 (6): 1509–17. doi:10.1113/jphysiol.2007.150029. PMC 2375708. PMID 18202093.
- ↑ 15.0 15.1 Perycz, M.; Urbanska, A. S.; Krawczyk, P. S.; Parobczak, K.; Jaworski, J. (2011). "Zipcode Binding Protein 1 Regulates the Development of Dendritic Arbors in Hippocampal Neurons" (PDF). Journal of Neuroscience. 31 (14): 5271–5285. doi:10.1523/JNEUROSCI.2387-10.2011. PMC 6622686. PMID 21471362. Archived (PDF) from the original on 2017-09-22.
- ↑ 16.0 16.1 Kandel, Eric R. (2003). Principles of neural science. (4th ed.). Cambridge: McGrawHill. ISBN 0-8385-7701-6. https://archive.org/details/isbn_9780838577011.
- ↑ 17.0 17.1 Koch, Christof (1999). Biophysics of computation : information processing in single neurons. New York [u.a.]: Oxford Univ. Press. ISBN 0-19-510491-9.
- ↑ 18.0 18.1 Häusser, Michael (2008). Dendrites (2nd ed.). Oxford: Oxford University Press. ISBN 978-0-19-856656-4.
- ↑ 19.0 19.1 Barnett, MW; Larkman, PM (Jun 2007). "The action potential". Practical Neurology. 7 (3): 192–7. PMID 17515599.
References
- Lorenzo, L. E.; Russier, M; Barbe, A; Fritschy, J. M.; Bras, H (2007). "Differential organization of gamma-aminobutyric acid type a and glycine receptors in the somatic and dendritic compartments of rat abducens motoneurons". The Journal of Comparative Neurology. 504 (2): 112–26. doi:10.1002/cne.21442. PMID 17626281. S2CID 26123520.
References
External links
- 模板:OklahomaHistology - "Slide 3 Spinal cord"
- Dendritic Tree - Cell Centered Database
- Stereo images of dendritic trees in Kryptopterus electroreceptor organs
- - "Slide 3 Spinal cord"
- Dendritic Tree - Cell Centered Database
- Stereo images of dendritic trees in Kryptopterus electroreceptor organs
= = = 外部链接 = = = =-”幻灯片3脊髓”
- 树突树细胞为中心的数据库
- 氪星电感器器官中树突树的立体图像
Category:Neurohistology Category:Neuroplasticity
类别: 神经组织学类别: 神经可塑性
This page was moved from wikipedia:en:Dendrite. Its edit history can be viewed at 树突/edithistory