异速生长律 Allometry
引言
近年来,随着所谓的“大数据”时代来临,复杂系统科学研究的基本方法论正在发生着天翻地覆的变化。从早期的纯哲学思辨,再到后来的计算机模拟,复杂系统研究终于开始采用“物理学范式”:从实证数据中来,得到一般性理论,再到具体的预测和实验中去。
物理学范式的一个成功的应用案例就是万有引力的研究(记得王雄曾经做过一次关于引力理论的讲座)。的确,从第谷、伽利略再到牛顿与爱因斯坦,物理学家花费了将近300年的时间,终于将行星运动的观测数据与完美的数学理论:爱因斯坦广义相对论联系了起来。这就是物理学的范式,一步一步地,虽然举步维艰,但是终于达到了可以精确预测星体运行的物理学。
在复杂系统研究领域,随着人们更加自觉地开始利用大量的数据,一些类似伽利略行星三大定律的定量规律已经慢慢地浮现出来。而其中最显眼的一条普适规律就是异速生长律了。所谓的异速生长律,就是指系统的两个宏观变量X和Y之间服从一个幂律方程:
[math]\displaystyle{ Y=c X^{\alpha} }[/math]
其中[math]\displaystyle{ c }[/math]和[math]\displaystyle{ \alpha }[/math]都是常数。这个方程有其深刻的内涵,目前已被证实适用于从细胞到国家横跨将近三十个数量级的复杂系统。
生物学中的异速生长律
异速生长 Allometry一词的来源
异速生长Allometry是关于身体大小与形状[1]、解剖学、生理学以及人类行为[2]间关系的研究,于1892年被Otto Snell[3],1917年被D'Arcy Thompson[4]和1932年被朱利安·赫胥黎 Julian Sorell Huxley[5]首先阐述。异速生长是个著名的研究论题,尤其在统计形状分析及活机体不同部分生长率研究的生物学上。应用之一就是在各种昆虫种类研究上,即整个身体中的某个小部位的变化,能够导致四肢附件如腿、触角等呈现巨大的不成比例的变化。生物学家Huxley 和Tessier[6] 1936年通过研究招潮蟹(fiddler crab)的钳子大小随着螃蟹身体而变化,发现了两者之间的幂律依赖关系,如下图所示:

随着招潮蟹从小长到大,它的身体在某个时刻t的尺寸(用螃蟹壳的宽度来衡量)是X(t),而它的钳子的大小(如上图左图红色线段所示)用Y(t)来表示,则这两个变量之间服从下面的方程:
[math]\displaystyle{ Y(t)=c X(t)^{\alpha} }[/math]
在这里[math]\displaystyle{ c }[/math]和[math]\displaystyle{ \alpha }[/math]都是常数。如果对上面等式两边取对数,则得到:
[math]\displaystyle{ \log(Y(t))=\log(c)+\alpha \log(X(t)) }[/math]
可以看出,[math]\displaystyle{ \log(Y(t)) }[/math]和[math]\displaystyle{ \log(X(t)) }[/math]构成了一个对线性依赖的变量。估计[math]\displaystyle{ a }[/math]指数可从类型2回归,如主轴回归或压轴回归分析来解释两变量的变化,与线性回归和最小二乘法回归相反,它不解释自变量(如对数体重)的误差方差。其他方法包括测量误差模型和特别类型的主成分分析法。异速生长常研究物种大小比率方面的形状差异。不同大小但共同形状的两个物种会在某个比率上有其共有维度。例如,一个生物体随年龄生长大小发生变化但形状却一直相似。个体发育学异速生长研究常用蜥蜴或蛇作为模式体。因为他们出生后都没有双亲育幼并且幼年与成年身体大小相差很大。蜥蜴在个体发育史上展现出明显的异速生长变化。除了关注生长,异速生长还研究给定年龄(或性别)下个体的形状变化,被称为静态异速生长。而物种间的比较研究常用于探索种间或进化的异速生长(另见系统发育比较方法)。
通过将实际测量的螃蟹尺寸数据画在双对数坐标轴上,并且用最小二乘法(Ordinary Linear Regression)可以从实际数据估计出参数[math]\displaystyle{ c }[/math]和[math]\displaystyle{ \alpha }[/math](如上图右图所示)。结果他们发现,实际估计出来的参数[math]\displaystyle{ \alpha }[/math]大于1(大概为1.57,为了对比,斜率为1的直线在上面右图所示),这就意味着如果招潮蟹体积增大了2倍,它的钳子的体积增长倍数要大于2倍,因此钳子的增长速度要远快于它的体积增长速度,这就是异速生长(Allometry或者Allometric scaling)一词的来源。(参考[7])
Allometry是一个合成词,allo-表示相异的,而-metry这个后缀则有测量、度量的意思。因此,这两个词根合起来就是相异的两种测量。而这里的相异又是何含义呢?我们要先理解Allometry的反义词Isometry一词,这里Iso是表示相同的。在上面的例子中,如果螃蟹钳子的生长速度和它的身体的生长速度是相同的,也就是说幂律指数[math]\displaystyle{ \alpha=1 }[/math],那就表示钳子和它的体积是线性的,它们以相同的速度增长(Isometry)。但是,事实上,我们测量到的真实数据告诉我们,这两个变量的生长速度并不一致,钳子快,身体慢,它们两者不协调,因此,就称为异速生长。
根据Wikipedia上面的解释,这里所说的同速增长Isometry并不一定就等于是线性增长关系,而是一种正常的按照欧几里德几何的增长方式。因此,Isometry就成为了一种作为参考和比较的类似于统计上的零模型的东西(Null model)。
比如说,对于欧几里德几何体来说,一个物体的面积的增长与该物体体积的增长呈现出2/3的幂律关系,即面积正比于体积的2/3次幂。让我们考虑生物体,例如人类大腿的截面面积,假如随着人体积的增长,截面面积呈现了2/3幂次的增长,那么该截面面积就是同速生长(相比较欧几里德几何体来说)。但是如果增长速度不等于2/3,那么我们就说大腿截面面积的增长属于异速生长。
等距生长和几何相似性
等距(缩放)生长是指生物体大小在生长过程或进化过程中呈比例关系变化。这样的例子在青蛙上被发现。除了青蛙变态后几周里的很短时间外,青蛙就是等距缩生长的[8] 。因此,青蛙的腿总是与其身体呈正比例变化,即使青蛙自身大小显著增加时也是如此。
等距缩放由平方-立方律决定,一个机体如果在长度上等距缩放地成倍增长,则其表面积将会增长4倍,而其体积和体重将会增长8倍。这时机体就会产生问题:机体要有8倍的活组织需要去支持,但呼吸器官的表面积却只增长了4倍,就会造成大小尺度与机体需求的不匹配。同样地,机体的腿部需要支撑起增长了8倍的体重,可它骨头和肌肉强度所依赖的断面积,却也只增长了4倍。因此,这样的假设性的机体需要经历骨头和肌肉负载变小两倍,或者是通过异速生长来避免这样的情况发生。
在尺度研究中,等距缩放常被用作零假设。即作为证明是迫使出现异速生长而偏离等距性的生理学证据。
任何非等距生长变化就是异速生长,伽俐略在其《关于两个新科学的对话》中讨论的典型例子就是关于兽类骨骼,即其骨骼因身体尺寸增加而会变得更强健和增大。异速生长常从以体重、体长等因素为基础的缩放标度幂指数来表示。典型的异速生长可能出现的情况是,所有体积相关的特征值随体重呈比例变化,所有面积相关的特征指数依体重2/3次方变化,而所有长度相关的特征依体重1/3次方变化。统计分析后表明,如果一个体积相关的特征相对体重缩放达到0.9次方,那么就称为“负向异速生长”,这时值会小于等距缩放的预测,相反地,如果一个面积相关的特征相对体重缩放到0.8次方,其值会高于等距缩放预测而机体可被称为“正向异速生长”。一个正向异速生长的例子是一些巨蜥属物种,其四肢相对要比相关的大体积物种长[9],对一些鱼类也是这样,
如何判定异速生长
去判断是否是等速还是异速生长,就需要去判定比较数据中变量间的相关关系,最重要是要判定数组中缩放关系是不是与期望的关系偏离(比如与等距生长相比)。应用的工具就是维度分析,用其来分析判定期望斜率是很有用的[10]。这个“期望”斜率,如果已知,对于判定异速生长就十分重要,因为缩放变量要与其他的进行比较,而这个是个衡量的尺子。比如说,如果只知道体重相对长度以斜率5进行缩放生长并无太大意义,除非你知道等距生长斜率是3,那才意味着体重在急剧增长。 例如,根据1950年Hill提出[11]和Wilson 2000[12]解释的几何相似性模型,不同大小的青蛙应该能够跳得一样远,但实际上大个青蛙当然能跳得更远。维度分析就是用以平衡这样的在公式中存在的差异,来判定期望斜率的。

如果尝试去找到体重与动物特征长度间关系的斜率,你可以以体重单位为Y轴,长度单位为X轴,由于体重与体积相当而体积是长度的立方,因此,在双对数图中你所期望的斜率就是3(log10(L3)/log10(L1)=3),这是一条直线的斜率,而实际情况下,大多数采集到的数据都不是自动接近直线的,因此要进行数据转换。比较数据时需要记住的是,比较的特征如头长对头宽可以与头长对体长的结果不同,不同的特征会不同的缩放率。
常用的转换数据的方法是做对数转换,转换两轴而执行线性回归。这可使数据正态化,使用线斜率分析趋势时更容易。分析之前你应该有一个预测斜率作为比较之用。数据转换与线性回归后,你可用95%可信度的最小二乘回归或降低主轴分析法。有时这两种方法会产生不同的结果,但大多数情况下是一致的。如果预测斜率在置信区间之外,则是异速生长,而如果假定动物以斜率5生长且有显著差异的值,那么与预测值相比,体重在这种动物上会生长非常快,这是正向异速生长。如果预测斜率是3,实际上某一有机体体重的缩放生长则为1(假定斜率也是显著差异的),那么这则是负向异速生长。
其他例子:力量是依赖肌肉横截面面积的,即L2,如果比较力量与长度,那么你的预测斜率是2。或者,这个分析也可以以乘方回归来完成。将你的数据间关系做图,拟合功率曲线,它会给你一个形式为y=Zx^number的公式,这个number是你数据点间的关系。这样做的缺点是做统计分析时会有些困难。
生理尺度生长
许多生理生化过程(心率、呼吸率、最大繁殖率等)都表现出尺度生长,且大多数与动物表面积及体重(体积)比率有关。个体代谢率也受制于尺度生长影响。
代谢率与体重
针对动物自己的体重和基础代谢率作图,可得到对数化的一条直线,显示出幂定律关系。一般来说动物基础代谢率表现出负异速生长,即如克萊伯定律所述,基础代谢率的尺度变化与体重的0.75次幂成正比。这就意味着,大个体动物(如象)有较低的单位代谢率真和心率,相比小个体动物(如鼠)而言,这条直线被称为“鼠像曲线”。
Max Kleiber 提出了下列异速生长方程来表述基础代谢率与动物体重间关系[13],统计分析中其截距为70而斜率为0.75:
- [math]\displaystyle{ \mathrm{Metabolic\ Rate} = 70 M^{0.75} }[/math]
[math]\displaystyle{ M }[/math]为体重,代谢率以千卡/天计。
克莱伯定律像是动物循环系统物理学和几何学特性的结果。同一物种中,年轻的个体比老的个体每份重量所占的呼吸更多,因为它们体重中中组织结构的比例更大,而储能的比例较小。结构质量需要耗费养护能量,而储能的质量则不需要。
基于物种的多样性,异速生长关系并非都是线性的或是对数式的,例如,哺乳类的最大奔跑速度与体重间就表现出极复杂的关系,最快的短跑者反而是中等体型者。
异速生长的肌肉特征
动物的肌肉特征在大多数不同大小的动物中均相似,尽管肌肉大小与形状常因环境限制因素而变化,肌肉组织可自身维持它的可伸缩性且并不依赖动物大小而变化。肌肉的生理生长会影响肌肉纤维的数量和它们的内在速度以决定动物运动效率与最大功效。肌肉重复的速度大概地与动物体重的立方根成反比例变化,比如麻雀飞行肌的内在频率与鹳的相比就是这样。
不同物种个体大小的决定因素
决定一个动物体体重与大小的因素很多,这些因素常常在进化尺度上影响着物种个体大小,但一些比如食物可利用性、生境大小等因素也能迅速作用于一个物种,具体如:
- 生理因素影响
- 基本的生理学设计在一个物种的大小方面扮演重要角色,比如,封闭血循系统的动物就要比开放血循系统的动物个体大。[13]
- 肌体机械因素影响
- 机械设计也能决定一个物种的最大可能大小,具有内骨骼的动物就倾向于比具外骨骼的动物个体大。[13]
- 生境因素影响
- 贯穿其进化史的生境对一种动物来说也是决定其大小的重要因素。在陆地上,在一定面积上的顶位物种的体重与其可用陆地面积呈正相关。[14]但是,在任意给定面积的陆地上,也有大量的“小”物种,因此,更多的决定因素可能来源于生态条件、进化因素、食物可利用性;大捕食者的小种群依赖于小猎物物种的大种群存活。在水生环境中,最大的动物能够产生并有比陆地动物大得多的体重,因为在水中不需要克服重力带来的限制。[11]
各种异速律
在上面的例子中,我们是将同一个生物个体不同时刻的钳子大小和个体大小两个变量作为数据点画在双对数坐标下得到异速生长律,这种异速生长律被称之为个体发生异速律(ontogenetic allometry )。我们也可以这样做:在同一个时间,找来一堆招潮蟹,这堆招潮蟹有大有小,我们将它们的钳子大小作为一个变量[math]\displaystyle{ Y_i }[/math](i表示个体),个头大小作为另一个变量[math]\displaystyle{ X_i }[/math],然后将这些数据点画在同一个坐标轴上。这个时候,我们仍然可能得到一个[math]\displaystyle{ Y_i }[/math]与[math]\displaystyle{ X_i }[/math]之间的幂律关系,但是这个幂律关系描述的是不同个体在同一个时刻的异速生长律。在生物学中,这被称为静态的异速律(Static allometry)。
另外,除了研究同一个物种不同的个体在同一个时间截面内的异速律,我们还可以研究不同的物种在同一个时间截面内平均值的异速生长律。前者称为种内的异速生长律(intraspecific allometry),后者称为种间的异速生长律(interspecific allometry)。
例如,我们考虑生物体的新陈代谢F和它们的重量M之间的异速生长关系。Max Rubner曾经对狗这个物种研究过这两个变量之间的关系,由于狗的品种繁多,而且他们的重量M的变化范围很大,因此画在双对数坐标下,会有一个很宽广的范围,可以研究种内的异速生长律。他的数据指出,狗种内的异速生长律的幂指数是2/3左右。后来又有人用鸟来做种内的异速生长律,结果也接近2/3。
然而,Kleiber(1932)发现,如果我们研究种间的F与M之间的关系。即找来不同的物种,并测量每个物种多个个体的平均新陈代谢量以及生物体重量,然后将这些数据点画在同一张图上,就会得到幂指数为3/4的异速生长律。因此,我们看到尽管是同一对变量,种间和种内的异速生长律幂指数也是很不同的。
3/4幂律——生物中的“开普勒”定律
尽管当前生物学已经积累了超海量数据,定量化研究也是势在必行。但是笔者认为,在众多的定量规律之中,Kleiber预言的生物体的新陈代谢和生物量之间的3/4幂律是最重要的一个,也是最接近天体中的开普勒的候选者。
因为Kleiber的3/4定律不仅仅给出了所有不同物种的统一的幂律关系,还指出这些幂律关系具有完全同样的幂指数3/4,更让人吃惊的是,这个定律的适用范围非常广泛,小到分子、细胞器、细胞,大到鲸鱼、大象,新陈代谢与生物量之间的3/4次幂律始终成立。大自然再一次在看似纷乱复杂的生物世界中展示了它的统一的数学之美。
继Kleiber等在早期针对哺乳动物得到了3/4幂律之后,Geoffery West等人又将这一定律的适用范围进一步扩大到了微观生物世界。在2002年的PNAS刊物上的一篇文章《Allometric scaling of metabolic rate from molecules and mitochondria to cells and mammals》中[15]他们指出从完整的生物体,到细胞、线粒体甚至到色素细胞氧化酶分子,在这横跨了27个数量级的上,所有这些生命现象都遵从同一个指数的3/4幂律。如下图所示:
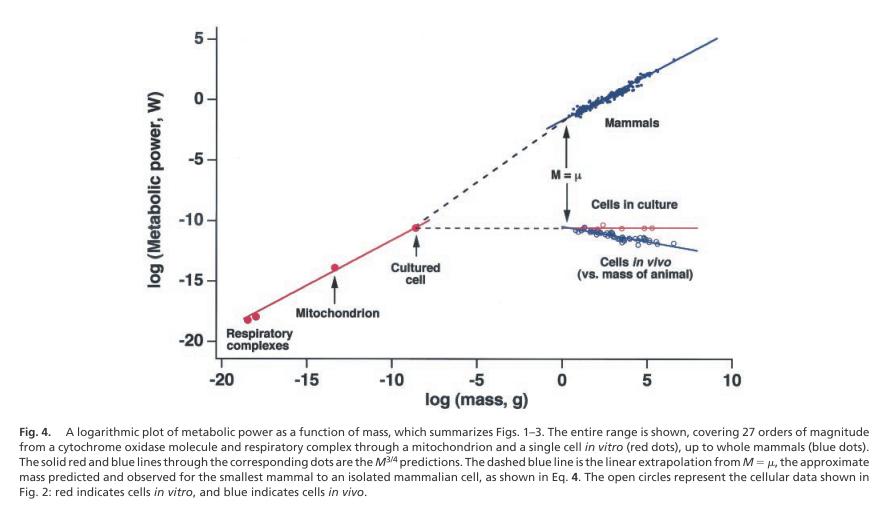

本图摘自:Allometric scaling of metabolic rate from molecules and mitochondria to cells and mammals
值得注意的是,对于不同尺度的生物系统来说,虽然他们的幂指数都非常接近3/4,但是这些直线的截距(也就是log(c)却不尽相同)。这意味着:处于同一条直线上的生物体具有相类似的网络结构(它们是同一种系统经过尺度变化,不断扩大出来的,这类似于一个个体从小到大的诚征)。而处在不同直线段上的生物体却不一定具有相似的结构。有关Kleiber律以及其它的种间的异速生长律的详细介绍,请大家参看:《流的探索》。
总而言之,我们可以将各种异速律进行如下的分类:
- 个体发生异速律(ontogenetic allometry )
- 静态异速律(Static allometry)
- 种内异速律(Intraspecific allometry)
- 种间异速律(Interspecific allometry)
从城市到国家
城市——创新的来源
异速生长现象如果说仅适用于生物系统,那么它还不够普适。但事实上,近期越来越多的研究表明,异速生长方程:[math]\displaystyle{ Y=c X^{\alpha} }[/math]是一个相当普遍的方程,只不过在其他系统中,较难以找到统一的幂指数a。
例如,当研究人类城市的时候,就会发现很多变量都随着城市规模(人口数)的增长而呈现异速生长方程。而这些变量所对应的幂指数大小不一。例如,在Luis Bettencourt等人的文章《Growth, innovation, scaling, and the pace of life in cities》中[16],作者研究了多种变量与城市人口之间的静态异速生长关系(同一个时间截面,不同的城市):

如上表所示,最左边一列是城市的各种指标(即变量Y),例如一个城市在一年内新申请的专利数量、城市的总产值(GDP)、总的房屋数目、总电量消耗、加油站数目等等。第二列则给出了异速生长方程Y=c X^a中的幂律指数。第三列是拟合优度R^2、第四列是在幂律方程中拟合点的数目,最后一列则指出了不同的数据来源,它们来自美国、中国、德国、欧洲的不同时期。
我们看到,这些变量的幂指数大小虽然都不一样,但是基本都可以分成三大类:>1,<1和约等于1。这三大类变量依次被称为超线性增长、亚线性增长和线性增长。幂律指数a的大小反映出了该变量随着城市的增长所体现出来的规模效应。
从幂律方程[math]\displaystyle{ Y=c X^{\alpha} }[/math]中变形,可以得到:
[math]\displaystyle{ Y/X=c X^{\alpha-1} }[/math]
在这里,方程的左边是人均拥有的Y变量(例如人均GDP,人均加油站数量等等)。如果a>1,那么方程右侧的幂指数就大于0,也就是人均拥有的Y变量会随着城市的增长而增大。如果a<1,则体现为人均Y会随着城市的增长而减小。如果a~1,就意味着人均Y不随城市规模而变化。
这用GDP作为Y来说明该方程的意义。GDP衡量的是一个经济单位在一段时期内创造的总产值。那么[math]\displaystyle{ Y/X }[/math]就表示一个城市的人均GDP。通过实证研究得到不同国家的城市的幂律指数都>1(在上表中基本上是1.2左右),也就意味着人均GDP会随着城市规模的增长而增长。
在中国,每年都会有大量的人口涌进像北京、上海这样的大城市,在全世界,超过一半的人口是居住在城市的,而且这种比例还在增大。为什么会出现这种情况?人们为何希望聚集到大城市呢?通过方程[math]\displaystyle{ Y/X=c X^{0.2} }[/math]不难看到,这是因为城市越大,人均产出(或者等价地即为人均收入)会随着城市规模的增大而增加。所以,城市越大就意味着平均来说每个人的收入就会增加(暂时忽略贫富之间的差距),所以很多人更愿意居住在大城市。
再来看一个变量Y为石油销售量。在表中,这个变量的幂指数a<1,这意味着随着城市的增大,人均所消耗的石油在减少。因此,人们聚集在一起形成大城市实际上是一种更加经济地使用石油资源的方式。
总而言之,根据该表格,很多与创新、财富增长有关的变量都具有超线性关系,而与能量、资源消耗有关的变量都呈现出亚线性关系,这些都说明了城市是一种具备规模经济效应的有机体。当然,城市规模的增加也有负面效应,例如,人均犯罪数量、人均艾滋病感染数量也都会随着城市规模的增加而增加。
国家的异速生长
在更大的国家尺度上来探索异速生长律。以下讨论主要基于张江已发表的一篇论文[17]。
研究异速生长定律,需要实证数据。张江发现,Mathematica 6.0软件之中就包含了大量的数据,且通过Mathamtica中CountryData命令,可以将自己的PC机与Wolfram公司提供的在线数据库相连,从而访问世界各国各方面的数据(包括地理、人口、经济、贸易等)。基于该数据库,张江展开了大量的关于异速生长律的研究。
在这个研究中,张的团队对273个国家或地区的104个数值型的变量进行了系统性的静态异速生长律研究。首先,将所有变量划分成了两类:1、广延型的变量;2、强度型变量。在物理中,所谓的广延型变量是诸如能量、质量、熵等具有子系统可加性的变量。在国家中,一些宏观总量,如人口、GDP、面积、出口、进口等都属于广延型变量。而平均的人均GDP,人口的生育率、死亡率等类似于物理中的温度、压强被称为强度量。
对于广延性的变量,张分别研究了它们和三种不同的国家尺度变量(分别是地理尺度,即国土面积、人口尺度,即总人口和经济尺度即GDP)做异速生长律分析,即探索Y=cX^a是否成立,这里X是面积、人口和GDP这三种不同的尺度。Y是各种广延性变量。
得到了一些有趣的结论如下:
1、国家作为更大的经济体与城市呈现出明显的差异
例如,如果仍然研究人口和GDP这两个变量的异速生长关系,就会发现虽然[math]\displaystyle{ Y=c X^a }[/math]这个方程仍然成立,但是幂指数a会非常接近于1,而不是大于1,也就是说,对于国家来说,GDP和人口近似呈现出线性的关系。如下图:

在这张图中,每个数据点都表示一个国家。红色的直线和黑色的虚线分别是用普通最小二乘法和主轴回归法得到的对异速生长关系幂指数的估计值。在本研究中,主要采纳主轴法得到的结果,呈现的结果就是国家的人口与GDP之间的关系呈现线性。
也许你会认为这个数据并不好,噪声很大,因此统计的显著性并不高([math]\displaystyle{ R^2=0.59 }[/math]),这样得到的幂指数a的估计会不会不准呢?事实上,可以通过选择不同的属性来得到更加准确、显著性更高的人口与产出之间的关系,如下图:

通过把横轴的人口属性用就业的劳动力总数来替代,纵轴用等价购买力重新核算的GDP来代替普通的GDP,就会得到统计显著性更高的异速生长关系(事实上[math]\displaystyle{ R^2 }[/math]从0.59提高到了0.88),而幂指数始终在1附近,几乎没变。由于世界上不同国家的差异非常大,因此用等价购买力折算的GDP比直接用GDP会更能反映出国家的实际产出能力。而在人口方面,由于并不是所有的人口都处于就业而产生产出的状态,因此利用就业的劳动力会去除更多的噪音。由此得到结论就是对于国家来说,GDP与人口成正比。
由此,已经得到了一个与城市截然不同的结论。在城市中,人口与GDP的关系是超线性的,即随着城市规模的增大,人均产出会增加。但是对于国家来说,人口越多的国家并不意味着更富裕,即越高的人均GDP(想一想印度和中国这两个国家,在图中的最右侧的两个点,就可以知道了)。事实上,在Bettencourt的城市文章[16]进行了对比发现了更多的不同:

从这张表,可以清楚地知道,随着人口的上升,城市的人均使用面积会减少,人均产出、电力消耗、感染艾滋病的总数会提高,石油消耗会减少,这体现了城市的规模效应。对于国家来说,随着人口的增加,人均用地、电力消耗、石油消耗、艾滋病感染都会增加,而人均产出近似不变,这说明,相比较城市来说,国家这种更大规模的系统并不呈现出规模经济效应。
为什么会存在这样的不同呢?很有可能国家相对于城市来说,他的封闭性越差,人口、物品、能量的流入和流出会受到更大的限制。这导致了国家不能形成有效的自组织系统。另外,由于城市属于人口密集的地区,而国家中大片的土地是处于没有人口居住的农村地区,所以国家的总产出近似与人口成正比而并非呈现出超线性关系。
GDP会比人口更好地衡量国家的尺度
张通过研究发现,很多宏观变量都与国家的GDP而非人口形成了显著性很高的幂律关系,如下表所示:

可以看到,从石油消耗、电力消耗到出口总额、进口总额,再到电话线长度、收音机电台个数、互联网用户数、公路总长度都可以用国家的GDP得到预测。而且,统计的显著性R^2都高于0.6。
有几个幂律关系非常有趣,例如进口总额、出口总额与GDP的关系。如果把国家看作是由货币流构成的宏观系统,那么进口总额、出口总额类似于国家经济体的心尘代谢,而GDP相当于经济体的生物体的存量。因此,出口、入口与GDP的异速生长律可以看作是经济流的Kleiber律。
另外一个有趣的幂律关系是碳排放(Carbon Emission)与GDP之间的超线性关系。根据研究,碳排放是与GDP呈现显著性很高的超线性关系的,因此GDP越高,碳排放就会越高。这是一种系统性的规律,很难通过个体行为而改变。因此,要抑制更多的碳排放,恐怕唯一的办法就是减少人类的产出。
最后,张还研究了各种强度量与人均GDP之间的关系。如下图展示的两个例子:

随着国家变得越来越富有(人均GDP的提高),总和生育率下降而平均年龄生高。而且这些强度量与人均GDP呈现了幂律的关系。张的团队猜想这类幂律关系与人类作为生物体需要进行新陈代谢,因此必然遵循代谢生态学的约束(参看《流的探索》)。
标度律现象
近年来,人们还发现了普遍的标度律现象(Scaling)。所谓的标度现象就是大家俗称的幂律现象,
虽然这些实证规律都称为标度律(或者幂律),但是与异速生长律有着重要的区别。一个明显的区别就是,异速生长律关注的是两个变量之间的幂律关系,即:
[math]\displaystyle{ Y\sim X^{\alpha} }[/math]
而其它的标度律则关心的是单一一个个变量的幂律分布。例如Pareto定律就在说对于经济系统中的高收入人群来说,他们的收入X服从幂律分布,即你发现收入在区间[math]\displaystyle{ [x,x+dx] }[/math]之间的人的比例是[math]\displaystyle{ x^{-a} }[/math]。即:
[math]\displaystyle{ p(x)\sim x^{-a} }[/math]
注意这里只有收入X是研究的变量,而[math]\displaystyle{ p(x) }[/math]是刻画变量X分布性质的概率密度函数,所以它是单一变量的幂律。
2012年2月,生物学家M. Stumpf和数学家M.A. Poter在Science上撰写《Critical Truths About Power Law》[18]一文声称很多幂律分布现象(例如复杂网络中的无标度分布)都不具有统计上的严格性,而且其生成机理也缺乏一定的深刻性。相反,异速生长标度律现象则统计意义上更加严格而且其机理也是很深刻的。
对于很多实证数据来说,单变量的分布往往不是严格的幂律分布,一般多呈现为头部偏向于指数或者对数正态分布,而尾部的一个小区域才会出现幂律分布(即常说的幂律尾现象)。而且很有可能这些数据的真实分布服从稳定分布而非严格的幂律分布(参见科普文章:《稳定分布与广义中心极限定理》[19])。相比较来说,双变量幂律也就是今天重点要说的异速生长法才更接近开普勒定律的地位。
总结来看,从细胞到国家,无论是生物系统、生态系统还是人类经济社会系统都遵从相同的幂律方程:
[math]\displaystyle{ Y=cX^{\alpha} }[/math]
这就是异速生长律,它是一种横贯了不同复杂系统的精确而定量的数学方程。
相关阅读
参考文献
- ↑ Small, Christopher G. (1996). The Statistical Theory of Shape. Springer. p. 4. ISBN 0-387-94729-9.
- ↑ Damuth J (February 2001). "Scaling of growth: plants and animals are not so different". Proc. Natl. Acad. Sci. U.S.A. 98 (5): 2113–4. Bibcode:2001PNAS...98.2113D. doi:10.1073/pnas.051011198. PMC 33381. PMID 11226197.
- ↑ Otto Snell (1892). "Die Abhängigkeit des Hirngewichts von dem Körpergewicht und den geistigen Fähigkeiten". Arch. Psychiatr. 23 (2): 436–446. doi:10.1007/BF01843462.
- ↑ Thompson, D'Arcy W (1992). On Growth and Form (Canto ed.). Cambridge University Press. ISBN 978-0-521-43776-9. http://books.google.com/books?id=7_F4OUJmLFcC.
- ↑ Huxley, Julian S. (1972). Problems of Relative Growth (2nd ed.). New York: Dover. ISBN 0-486-61114-0.
- ↑ [1],文章:Allometry
- ↑ [2], Nature文章:Allometry: the study of biological scaling
- ↑ Emerson SB (September 1978). "Allometry and Jumping in Frogs: Helping the Twain to Meet". Evolution. 32 (3): 551–564.
- ↑ Christian, A.; Garland, T., Jr. (1996). "Scaling of limb proportions in monitor lizards (Squamata: Varanidae)" (PDF). Journal of Herpetology. 30 (2): 219–230.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Pennycuick, Colin J. (1992). Newton Rules Biology. Oxford University Press. p. 111. ISBN 0-19-854021-3.
- ↑ 11.0 11.1 Hill, A.V. (April 1950). "The dimensions of animals and their muscular dynamics". Science Progress. 150 (4176): 820. doi:10.1038/164820b0.
- ↑ Wilson RS, Franklin CE, James RS (June 2000). "Allometric scaling relationships of jumping performance in the striped marsh frog Limnodynastes peronii". J. Exp. Biol. 203 (Pt 12): 1937–46. PMID 10821750.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ 13.0 13.1 13.2 Willmer, Pat (2009). Environmental Physiology of Animals. Wiley-Blackwell.
- ↑ Burness, G. P.; Diamond, Jared; Flannery, Timothy (2001). "Dinosaurs, dragons, and dwarfs: The evolution of maximal body size". Proc. Natl. Acad. Sci. U.S.A. 98 (25): 14518–23. Bibcode:2001PNAS...9814518B. doi:10.1073/pnas.251548698. PMC 64714. PMID 11724953.
- ↑ West, Geoffery (2002). "Allometric scaling of metabolic rate from molecules and mitochondria to cells and mammals". PNAS. 99: 2473–2478.
- ↑ 16.0 16.1 Luís, M. A. Bettencourt* José Lobo, Dirk Helbing, Christian Kühnert, Geoffrey B. West (2007). "Growth, innovation, scaling, and the pace of life in cities". PNAS. 104 (17): 7301–7306.
- ↑ Zhang, Jiang. "Allometric scaling of countries". Physica A. Event occurs at 2010: 4887–4896.
- ↑ Stumpf1, Michael P. H. "Critical Truths About Power Law". Science. Event occurs at 2012: 665-666.
- ↑ [3],稳定分布与广义中心极限定理
延伸阅读
- Calder, W. A. (1984). Size, function and life history. Harvard University Press. ISBN 0-674-81070-8.
- McMahon, T. A.; Bonner, J. T. (1983). On Size and Life. Scientific American Library. ISBN 0-7167-5000-7.
- Niklas, K. J. (1994). Plant allometry: The scaling of form and process. University of Chicago Press. ISBN 0-226-58081-4. http://books.google.com/books?id=2th19CVNWtcC.
- Peters, R. H. (1983). The ecological implications of body size. Cambridge University Press. ISBN 0-521-28886-X. http://books.google.com/books?id=OYVxiZgTXWsC.
- Reiss, M. J. (1989). The allometry of growth and reproduction. Cambridge University Press. ISBN 0-521-42358-9. http://books.google.com/books?id=SkguxvCDDFcC.
- Schmidt-Nielsen, K. (1984). Scaling: why is animal size so important?. Cambridge: Cambridge University Press. ISBN 0-521-31987-0. http://books.google.com/books?id=8WkjD3L_avQC.
- Samaras, Thomas T. (2007). Human body size and the laws of scaling: physiological, performance, growth, longevity and ecological ramifications. Nova Publishers. ISBN 1-60021-408-8. http://books.google.com/books?id=PCU0RwDI6c4C.
本中文词条由用户:打豆豆、用户:李媛翯参与编译,欢迎在讨论页面留言。
本词条内容源自wikipedia及公开资料,遵守 CC3.0协议。